You might also like
- Purification and Properties of L-Glutaminase-L-Asparaginase From Pseudomonas AcidovoransDocument8 pagesPurification and Properties of L-Glutaminase-L-Asparaginase From Pseudomonas AcidovoransPrasanna KumarNo ratings yet
- Inhibition of Aldehyde Dehydrogenase IN Brain and Liver by CyanamideDocument5 pagesInhibition of Aldehyde Dehydrogenase IN Brain and Liver by CyanamideCristina Martín JiménezNo ratings yet
- 1981 Hizukuri, Takjzda, Yasuda 1981Document9 pages1981 Hizukuri, Takjzda, Yasuda 1981Łukasz GraczykNo ratings yet
- Astaxanthin Formation by The Yeast Phafia RhodozymaDocument11 pagesAstaxanthin Formation by The Yeast Phafia RhodozymaFlorencia GorordoNo ratings yet
- Synthesis and Interconversion of Acids Developing Cotyledons of Pea (Pisum Sativum L.)Document5 pagesSynthesis and Interconversion of Acids Developing Cotyledons of Pea (Pisum Sativum L.)Siti Munawaroh ITN MalangNo ratings yet
- Plntphys00211 0097Document7 pagesPlntphys00211 0097yasmineat29No ratings yet
- 1977 CazzuloDocument3 pages1977 CazzuloKeyla MaydeeNo ratings yet
- 10 1007@BF00390820Document8 pages10 1007@BF00390820sayyed bassir ajellehNo ratings yet
- Glucosamine Metabolism: Journal or Bioloqical Chemistry Vol. 235, No.6, May 1960Document10 pagesGlucosamine Metabolism: Journal or Bioloqical Chemistry Vol. 235, No.6, May 1960Melin YohanaNo ratings yet
- SERAHDocument9 pagesSERAHLoveth BethelNo ratings yet
- Effect of DrugsDocument6 pagesEffect of DrugsMohammad AlhasanNo ratings yet
- Article:: Ii. Oxidation of Sugars by An Carbohydrate Metabolism of Citrus FruitsDocument7 pagesArticle:: Ii. Oxidation of Sugars by An Carbohydrate Metabolism of Citrus FruitsJeremia Kevin EmEmNo ratings yet
- 08 - Chapter 2Document14 pages08 - Chapter 2The FrequencyNo ratings yet
- Characterization AcetylDocument14 pagesCharacterization AcetylhtersadrNo ratings yet
- Prostaglandin HistoryDocument6 pagesProstaglandin HistoryRishi AllamNo ratings yet
- Isolation and partial characterisation of α-amylase components evolved during early wheat germinationDocument13 pagesIsolation and partial characterisation of α-amylase components evolved during early wheat germinationDwi Surya AtmajaNo ratings yet
- Sc. College Journal Res.72Document174 pagesSc. College Journal Res.72Pratik AmanNo ratings yet
- Dermorphin Related PeptidesDocument5 pagesDermorphin Related PeptidesKambo VivaNo ratings yet
- Application of RT-PCR For Indexing Avocado Sunblotch ViroidDocument4 pagesApplication of RT-PCR For Indexing Avocado Sunblotch ViroidErick RamirezNo ratings yet
- The Acyl-Enzyme Intermediate and The Kinetic Mechanism of The Glyceraldehyde 3-Phosphate Dehydrogenase ReactionDocument13 pagesThe Acyl-Enzyme Intermediate and The Kinetic Mechanism of The Glyceraldehyde 3-Phosphate Dehydrogenase ReactionLizi SorciaNo ratings yet
- Purification and Characterization of An Extracellular Agglutinin From Containing GlycoconjugatesDocument8 pagesPurification and Characterization of An Extracellular Agglutinin From Containing GlycoconjugatesBudi UtomoNo ratings yet
- The Amino Acid Sequences of Human and Pig L-Arginine:glycine AmidinotransferaseDocument7 pagesThe Amino Acid Sequences of Human and Pig L-Arginine:glycine AmidinotransferaseMona ChasdianaNo ratings yet
- 0065 2571 (72) 90019 2Document20 pages0065 2571 (72) 90019 2Muhammad Akbar SusenoNo ratings yet
- Fermentative Metabolism To Produce Hydrogen Gas and Organic Compounds in A CyanobacteriumDocument4 pagesFermentative Metabolism To Produce Hydrogen Gas and Organic Compounds in A Cyanobacteriumraisya farah monicaNo ratings yet
- Takeyoshiminaga1978 PDFDocument9 pagesTakeyoshiminaga1978 PDFAracely FloresNo ratings yet
- Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform ExtractionDocument4 pagesSingle-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform ExtractionAnggraeni Arum SNo ratings yet
- Brain Acetylcholinesterase As A N Detector of Organophosphorus A N D Carbamate Insecticides in WaterDocument6 pagesBrain Acetylcholinesterase As A N Detector of Organophosphorus A N D Carbamate Insecticides in WaterRodrigosqmNo ratings yet
- Production of Aldehyde Oxidases by Microorganisms and Their Enzymatic PropertiesDocument6 pagesProduction of Aldehyde Oxidases by Microorganisms and Their Enzymatic Propertiesapi-3743140No ratings yet
- Production of Lactase by Trichoderma SP.: Isil Seyis and Nilufer AksozDocument4 pagesProduction of Lactase by Trichoderma SP.: Isil Seyis and Nilufer AksozGuhan KANo ratings yet
- Purification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Document10 pagesPurification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Anastasya AmandaNo ratings yet
- Ja9b02611 Si 001Document23 pagesJa9b02611 Si 001Enziana MaharaniNo ratings yet
- Isolation and Characterization of E-64, A New Thiol Protease InhibitorDocument7 pagesIsolation and Characterization of E-64, A New Thiol Protease InhibitorHassanNo ratings yet
- Purification and Characterization of A Novel Solvent-Tolerant Lipase FromDocument4 pagesPurification and Characterization of A Novel Solvent-Tolerant Lipase Fromسید حسین عارفیNo ratings yet
- Purification and Characterisation of A Carboxylesterase From The Latex ofDocument16 pagesPurification and Characterisation of A Carboxylesterase From The Latex ofPol Melgar RuizNo ratings yet
- 10 1007@bf00522476Document12 pages10 1007@bf00522476thiagoNo ratings yet
- Spanac Si CoriandruDocument15 pagesSpanac Si CoriandruGaby IklprNo ratings yet
- Large Scale Production of D-Allose From D-Psicose Using Continuous Bioreactor and Separation SystemDocument5 pagesLarge Scale Production of D-Allose From D-Psicose Using Continuous Bioreactor and Separation SystemFabián Camilo OtáloraNo ratings yet
- Bonney 1979Document12 pagesBonney 1979Auliana FENo ratings yet
- Kinetic Characterization of Yeast Alcohol Dehydrogenases: J. Ganzhorn, David D, Hershey, Robert M, Gould, and BryceDocument8 pagesKinetic Characterization of Yeast Alcohol Dehydrogenases: J. Ganzhorn, David D, Hershey, Robert M, Gould, and BryceleztlyNo ratings yet
- Expression of The Saccharomyces Diastaticus Sta1 Gene in Brewing 1989Document6 pagesExpression of The Saccharomyces Diastaticus Sta1 Gene in Brewing 1989JefersonMatosdeColaresNo ratings yet
- Jurnal TekZImDocument10 pagesJurnal TekZImFestus Septian Z. YoesafatNo ratings yet
- Purification and Partial Characterization of Oxalate Oxidase From Leaves of Forage Sorghum (Sorghum Vulgare Var. KH-105) SeedlingsDocument5 pagesPurification and Partial Characterization of Oxalate Oxidase From Leaves of Forage Sorghum (Sorghum Vulgare Var. KH-105) SeedlingsAdy PrasetyoNo ratings yet
- Paper4 PDFDocument9 pagesPaper4 PDFJaimeParedesNo ratings yet
- 1 PBDocument5 pages1 PBCamilo Ernesto Araujo BarabasNo ratings yet
- Distribucion de Metilcytosina en La Piramide de Sec de Ac NucleicosDocument7 pagesDistribucion de Metilcytosina en La Piramide de Sec de Ac NucleicosJon Rojas PinoNo ratings yet
- Enzymology: Practical Manual BIOC231Document40 pagesEnzymology: Practical Manual BIOC231Shubham SutarNo ratings yet
- Reagents.: Materials and MethodsDocument6 pagesReagents.: Materials and Methodsedal_108No ratings yet
- Synthesis Characterization Oc-Adrenoceptors Parotid Slices Human MembranesDocument5 pagesSynthesis Characterization Oc-Adrenoceptors Parotid Slices Human MembranesLonkesNo ratings yet
- Plan Cience: Isolation and Properties of A Lectin From The Seeds ofDocument6 pagesPlan Cience: Isolation and Properties of A Lectin From The Seeds ofRana KameiNo ratings yet
- For Peer Review: Purification and functional characterization of an α-L-rhamnosidase from Penicillium citrinum MTCC-3565Document24 pagesFor Peer Review: Purification and functional characterization of an α-L-rhamnosidase from Penicillium citrinum MTCC-3565vinita_yadav23No ratings yet
- Pnas00658 0499Document3 pagesPnas00658 0499abril ibarraNo ratings yet
- Biochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumDocument4 pagesBiochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumPedro HamannNo ratings yet
- Zingiberene 2Document5 pagesZingiberene 2Luis MesenNo ratings yet
- Wang2004 Article Anti-oxidationOfAgarOligosacchDocument8 pagesWang2004 Article Anti-oxidationOfAgarOligosacchYasmin EkapratiwiNo ratings yet
- Sharath 03Document9 pagesSharath 03Dr. Sharath Kumar M NNo ratings yet
- J. Agric. Food Chem. 2012, 60, 7204 7210Document7 pagesJ. Agric. Food Chem. 2012, 60, 7204 7210Duong Pham QuangNo ratings yet
- Radial Diffusion Assay of NAD and NADP Dependent OxidoreductasesDocument3 pagesRadial Diffusion Assay of NAD and NADP Dependent OxidoreductasesNigel LoewNo ratings yet
- High Affinity of The Naturally-Occurring Biflavonoid, Amentoflavon, To Brain Benzodiazepine Receptors in VitroDocument3 pagesHigh Affinity of The Naturally-Occurring Biflavonoid, Amentoflavon, To Brain Benzodiazepine Receptors in VitroJuan Carlos LrNo ratings yet
- Gel Electrophoresis of ProteinsFrom EverandGel Electrophoresis of ProteinsMichael J DunnNo ratings yet
- Green 98 Glaucoma Arch OpthalmolDocument35 pagesGreen 98 Glaucoma Arch OpthalmolМатиас Себальос ГусманNo ratings yet
- Welch Et - Al 95 With Morphine JPharmExpTherDocument79 pagesWelch Et - Al 95 With Morphine JPharmExpTherМатиас Себальос ГусманNo ratings yet
- Hine Et - Al. 75, Blocks Abstinence, ScienceDocument10 pagesHine Et - Al. 75, Blocks Abstinence, ScienceМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument31 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument55 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument37 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- Hao. Food IntakeDocument31 pagesHao. Food IntakeМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument30 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument33 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument41 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманNo ratings yet
- Huitron-Resendiz 2001 PDFDocument40 pagesHuitron-Resendiz 2001 PDFМатиас Себальос ГусманNo ratings yet
- DePetrocellis Et - Al 98 Breast Cancer ProcNat'lAcadSci PDFDocument23 pagesDePetrocellis Et - Al 98 Breast Cancer ProcNat'lAcadSci PDFМатиас Себальос ГусманNo ratings yet
- 2008 Markle Et Rosch - Experiments of The Effects of Carbonization Oncultivated Plant SeedsDocument8 pages2008 Markle Et Rosch - Experiments of The Effects of Carbonization Oncultivated Plant SeedsLauraNo ratings yet
- Bio Test Assignment: There Are Two Parts of The Test Assignment. Part 1: Mcqs and Part 2: Essay Part 1: McqsDocument3 pagesBio Test Assignment: There Are Two Parts of The Test Assignment. Part 1: Mcqs and Part 2: Essay Part 1: Mcqsprmahajan18No ratings yet
- (Solvent) Diol Aldehyde-Alcohol Ketone-Alcohol Compound 1 Compound 2 Compound 3Document10 pages(Solvent) Diol Aldehyde-Alcohol Ketone-Alcohol Compound 1 Compound 2 Compound 3Ilias YacNo ratings yet
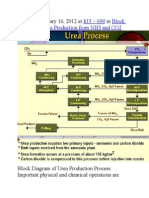
- Published January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2Document9 pagesPublished January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2himanshuchawla654No ratings yet
- Ncert Booster Test SeriesDocument23 pagesNcert Booster Test SeriesEr Sir100% (1)
- Biochem Viva Pool (1st Year MBBS)Document11 pagesBiochem Viva Pool (1st Year MBBS)TAHANo ratings yet
- Technical Data Sheet: SNDA-EAGLE (COCX35-97/60) 60%Document1 pageTechnical Data Sheet: SNDA-EAGLE (COCX35-97/60) 60%Fadi MagdyNo ratings yet
- Resistance Values of Structural and Finish Materials: Material R-Value Material R-ValueDocument1 pageResistance Values of Structural and Finish Materials: Material R-Value Material R-ValueRiyas UdheenNo ratings yet
- HW 1. StudentDocument3 pagesHW 1. Studentkitty2911No ratings yet
- BS en 17427-2022Document36 pagesBS en 17427-2022samir lahsaouiNo ratings yet
- Biomarkers For Geologists-A Practical Guide To The Application of Steranes and Triterpanes in Petroleum GeologyDocument90 pagesBiomarkers For Geologists-A Practical Guide To The Application of Steranes and Triterpanes in Petroleum GeologyReddy Setyawan67% (3)
- Pengaruh Pemberian Bahan Organik Terhadap PH, Kapasitas Tukar Kation (KTK) Dan C Organik Tanah Tukungan Pada Umur Yang BerbedaDocument5 pagesPengaruh Pemberian Bahan Organik Terhadap PH, Kapasitas Tukar Kation (KTK) Dan C Organik Tanah Tukungan Pada Umur Yang BerbedaMoza DarvinNo ratings yet
- Reaxys Anonymous 20131108 024356 919Document80 pagesReaxys Anonymous 20131108 024356 919ebi1364No ratings yet
- Module 1 - Chemical SafetyDocument21 pagesModule 1 - Chemical SafetyJason ErecillaNo ratings yet
- Exhanger Leakages in VDU - ModifiedDocument14 pagesExhanger Leakages in VDU - ModifiedJay LawsonNo ratings yet
- Al KanesDocument35 pagesAl Kanessimonatics08No ratings yet
- Ajac 2014022113435228Document6 pagesAjac 2014022113435228ajoilhamNo ratings yet
- Chemistry Investigatory Project Class 12Document17 pagesChemistry Investigatory Project Class 12Rosalin Acharya XIIC 15100% (1)
- Characterization of Volatile Compounds in Linden Blossoms: Tilia Curdata MillDocument6 pagesCharacterization of Volatile Compounds in Linden Blossoms: Tilia Curdata MillРусланNo ratings yet
- Biology 30 Unit C - Cell Division, Genetics, and Molecular Genetics - Chapter 18Document165 pagesBiology 30 Unit C - Cell Division, Genetics, and Molecular Genetics - Chapter 18kimmoNo ratings yet
- Control of Gel Time UprDocument6 pagesControl of Gel Time UprFredy HernándezNo ratings yet
- Phosphoric Acid PDFDocument76 pagesPhosphoric Acid PDFVeiliLookNo ratings yet
- Plastic From Potato StarchDocument22 pagesPlastic From Potato StarchMohammed EzzatNo ratings yet
- 2490-Article Text-5712-1-10-20200709Document8 pages2490-Article Text-5712-1-10-20200709Guadalupe Fernando Herrera AcostaNo ratings yet
- Reaktor 1Document1 pageReaktor 1fadliNo ratings yet
- Harga Jual ObatttttttttttttDocument12 pagesHarga Jual Obatttttttttttttsofia nofiantiNo ratings yet
- Esen Plastik A.S. GRP CatalogDocument50 pagesEsen Plastik A.S. GRP CatalogMarwan KhattabNo ratings yet
- حوض غدامسDocument5 pagesحوض غدامسMuhab Omar AbiedNo ratings yet
- Agro EmulsifiersDocument3 pagesAgro EmulsifiersFarizal HammiNo ratings yet
- T5. Polymeric Anti Redeposition (Clariant)Document16 pagesT5. Polymeric Anti Redeposition (Clariant)Maria Ofelia Toloza Vaca100% (1)