You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Biology Lesson Note Two For Grade 11.dox.Document2 pagesBiology Lesson Note Two For Grade 11.dox.Amanuel TesfayeNo ratings yet
- ExamDocument8 pagesExamfatiNo ratings yet
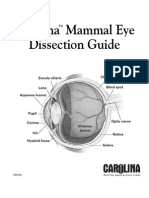
- Mammal Eye DissectionDocument4 pagesMammal Eye Dissectionnongnong2516No ratings yet
- V ZugastiAlvarez - Bird Island - Investigating BiodiversityDocument2 pagesV ZugastiAlvarez - Bird Island - Investigating BiodiversityVictoria ZugastiNo ratings yet
- Unit 7 - Evolution and Classification: Regents BiologyDocument24 pagesUnit 7 - Evolution and Classification: Regents BiologyTalijah JamesNo ratings yet
- EDNA Manual Eng v2 1 3bDocument94 pagesEDNA Manual Eng v2 1 3bNelly MarlinaNo ratings yet
- Holiday Homework Grade 12Document6 pagesHoliday Homework Grade 12RAUNAK DEYNo ratings yet
- Cell CycleDocument9 pagesCell CycleCrystal Joy RebelloNo ratings yet
- MC 102 Microbiology and ParasitologyDocument31 pagesMC 102 Microbiology and ParasitologyM StarkNo ratings yet
- Normas DRISDocument19 pagesNormas DRISRoger De Jesus Urrea LopezNo ratings yet
- OedogoniumDocument15 pagesOedogoniumDeepika KV100% (2)
- List of Subjects - SHS As of 2021Document2 pagesList of Subjects - SHS As of 2021Lavander BlushNo ratings yet
- Kellner Et Al. 2022 FaxinalipterusDocument32 pagesKellner Et Al. 2022 Faxinalipterusmauricio.garciaNo ratings yet
- Prep 4 Upsc: Test Series Topic Listing TEST 1-5Document4 pagesPrep 4 Upsc: Test Series Topic Listing TEST 1-5Babji ThothadiNo ratings yet
- Cell Cycle Report ProjectDocument17 pagesCell Cycle Report ProjectLawa KacakNo ratings yet
- 4 Basic Steps of Genetic Recombination EngineeringDocument1 page4 Basic Steps of Genetic Recombination Engineeringshainemary niones100% (1)
- Lesson Plan Earth and Life ScienceDocument3 pagesLesson Plan Earth and Life ScienceEmmanuel RonquilloNo ratings yet
- UntitledDocument55 pagesUntitledJi KimNo ratings yet
- Life Sciences Grade 10 Learner Support Document MAFUMANI SECONDARYDocument40 pagesLife Sciences Grade 10 Learner Support Document MAFUMANI SECONDARYdeveloping habit and lifestyle of praise and worshNo ratings yet
- Lecture 2 Biological MembraneDocument78 pagesLecture 2 Biological MembraneSyed Inam UllahNo ratings yet
- Textbook Bioinformatics Volume I Data Sequence Analysis and Evolution 2Nd Edition Jonathan M Keith Eds Ebook All Chapter PDFDocument53 pagesTextbook Bioinformatics Volume I Data Sequence Analysis and Evolution 2Nd Edition Jonathan M Keith Eds Ebook All Chapter PDFbrandy.arnold548100% (19)
- Microsyllabus BSC 301Document9 pagesMicrosyllabus BSC 301Anwita JhaNo ratings yet
- Language Development and AgeDocument304 pagesLanguage Development and AgejordicicNo ratings yet
- Research Paper AnxietyDocument7 pagesResearch Paper Anxietyeghkq0wf100% (1)
- Typological Design Strategy of FOA S ArchitectureDocument8 pagesTypological Design Strategy of FOA S ArchitecturePrzemyslawKolodziejNo ratings yet
- Flow Cytometry Capabilities GuideDocument16 pagesFlow Cytometry Capabilities GuideirmaNo ratings yet
- 3rd ObservationDocument44 pages3rd ObservationEnmar Petere-GuevarraNo ratings yet
- Thiesand Grossman 2006Document21 pagesThiesand Grossman 2006Be FahmiNo ratings yet
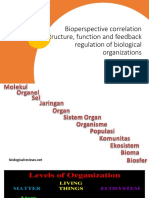
- (ENG) Bioperspektif Korelasi Struktur, Fungsi Dan Regulasi UmpanDocument28 pages(ENG) Bioperspektif Korelasi Struktur, Fungsi Dan Regulasi UmpanZuhrotul ZahrohNo ratings yet
- Art 4 25-35Document11 pagesArt 4 25-35Dicky AdipratamaNo ratings yet