You might also like
- Meth Impurity ProfilingDocument16 pagesMeth Impurity Profilingconker4No ratings yet
- Machine Learning Interviews - Lessons From Both Sides - FSDLDocument70 pagesMachine Learning Interviews - Lessons From Both Sides - FSDLneeraj12121100% (2)
- Getting Started Data Science GuideDocument76 pagesGetting Started Data Science GuideKunal LangerNo ratings yet
- AI in Indian Governance: Opportunities, Challenges and a Path ForwardDocument64 pagesAI in Indian Governance: Opportunities, Challenges and a Path Forwardneeraj12121No ratings yet
- Bioorganic and Medicinal ChemistryDocument19 pagesBioorganic and Medicinal ChemistryShefket DehariNo ratings yet
- God, Buber, GestaltDocument15 pagesGod, Buber, GestaltAriel Pollak100% (1)
- Ebook Getting Started With Design SystemsDocument24 pagesEbook Getting Started With Design SystemsJuan Carlos AcostaNo ratings yet
- Cot DLP FBS 3RD and 4THDocument11 pagesCot DLP FBS 3RD and 4THMherasul Pasaylo100% (2)
- Rishab Jain Internship GuideDocument10 pagesRishab Jain Internship GuideAdvik GuptaNo ratings yet
- 1-s2.0-S0960894X12014291-mainDocument5 pages1-s2.0-S0960894X12014291-mainhanNo ratings yet
- Determination Pyrazinamide in Human PlasmaDocument7 pagesDetermination Pyrazinamide in Human Plasmaervano1969No ratings yet
- Gjerde 2005Document9 pagesGjerde 2005Desi TrirahmaNo ratings yet
- Synthesis and in Vitro Biological Evaluation of Novel Pyrazole Derivatives As Potential Antitumor AgentsDocument10 pagesSynthesis and in Vitro Biological Evaluation of Novel Pyrazole Derivatives As Potential Antitumor AgentsAntônio Neto MachadoNo ratings yet
- In Vitro and in Vivo Evaluation of Novel Cinnamyl Sulfonamide Hydroxamate Derivative Against Colon AdenocarcinomaDocument14 pagesIn Vitro and in Vivo Evaluation of Novel Cinnamyl Sulfonamide Hydroxamate Derivative Against Colon AdenocarcinomajohnyeapNo ratings yet
- Analysis of Food Oligosaccharides Using MALDI-MS: Quantification of FructooligosaccharidesDocument9 pagesAnalysis of Food Oligosaccharides Using MALDI-MS: Quantification of FructooligosaccharidesseverithNo ratings yet
- Development and Validation of A LC/MS/MS Method For The Determination of Duloxetine in Human Plasma and Its Application To Pharmacokinetic StudyDocument14 pagesDevelopment and Validation of A LC/MS/MS Method For The Determination of Duloxetine in Human Plasma and Its Application To Pharmacokinetic StudyMohamed Medhat AliNo ratings yet
- Genotoxic Effect of - Hydroxy-4-Acetylaminobiphenyl On Human DNA: Implications in Bladder CancerDocument6 pagesGenotoxic Effect of - Hydroxy-4-Acetylaminobiphenyl On Human DNA: Implications in Bladder CancerSourashis BiswasNo ratings yet
- 2015 Article 570Document7 pages2015 Article 570Haile KassaNo ratings yet
- Research ArticleDocument13 pagesResearch ArticleMauro Porcel de PeraltaNo ratings yet
- QSAR AntikankerDocument8 pagesQSAR AntikankerHuda MuhammadNo ratings yet
- Isp 2Document5 pagesIsp 2wi bowoNo ratings yet
- High Performance Capillary Electrophoresis (HPCE) Determination of FR 900098, An Antimalarial Compound, in Aqueous Solutions and Human SerumDocument9 pagesHigh Performance Capillary Electrophoresis (HPCE) Determination of FR 900098, An Antimalarial Compound, in Aqueous Solutions and Human SerumВалерий СурнинNo ratings yet
- Abuse DrugDocument12 pagesAbuse DrugkorlNo ratings yet
- The In-Vitro Effect of Complementary and Alternative Medicines On Cytochrome P450 2C9 ActivityDocument8 pagesThe In-Vitro Effect of Complementary and Alternative Medicines On Cytochrome P450 2C9 ActivityNimra Naveed ShaikhNo ratings yet
- 10 1016@j Microc 2019 04 079Document8 pages10 1016@j Microc 2019 04 079Artem KulikovNo ratings yet
- Research ArticleDocument8 pagesResearch ArticleHyacinth A RotaNo ratings yet
- 365 Full PDFDocument8 pages365 Full PDFMiguel Machaca Flores (QuimioFarma)No ratings yet
- Dieterle 1977Document9 pagesDieterle 1977Carolina Calvache LunaNo ratings yet
- 10 1016@j Microc 2019 03 084Document8 pages10 1016@j Microc 2019 03 084Artem KulikovNo ratings yet
- Ajhg00066 0026Document12 pagesAjhg00066 0026ANELIZNo ratings yet
- Novel Hybrid Molecules of Isoxazole Chalcone Derivatives: Synthesis and Study of in Vitro Cytotoxic ActivitiesDocument7 pagesNovel Hybrid Molecules of Isoxazole Chalcone Derivatives: Synthesis and Study of in Vitro Cytotoxic ActivitiesRatnakaram Venkata NadhNo ratings yet
- S Imultaneous Determination of Neutral and Acidic Pharmaceuticals in WastewaterDocument9 pagesS Imultaneous Determination of Neutral and Acidic Pharmaceuticals in Wastewaterwatersoul.nNo ratings yet
- Detection of Human Urinary 5 Hydroxymethylcytosine by Stable Isotope Dilution HPLC-MS/MS AnalysisDocument7 pagesDetection of Human Urinary 5 Hydroxymethylcytosine by Stable Isotope Dilution HPLC-MS/MS Analysis承成蔡No ratings yet
- Tsuyoshi Minematsu, Jennifer Lee, Jiuhong Zha, Selina Moy, Donna Kowalski, Katsuyuki Hori, Koji Ishibashi, Takashi Usui, and Hidetaka KamimuraDocument11 pagesTsuyoshi Minematsu, Jennifer Lee, Jiuhong Zha, Selina Moy, Donna Kowalski, Katsuyuki Hori, Koji Ishibashi, Takashi Usui, and Hidetaka KamimuraHarry BalzacNo ratings yet
- Pharmacokinetics of Tramadol and Metabolites After Injective Administrations in DogsDocument6 pagesPharmacokinetics of Tramadol and Metabolites After Injective Administrations in DogsBoyITBNo ratings yet
- s11130 009 0111 620211016 31050 1u4wp1y With Cover Page v2Document9 pagess11130 009 0111 620211016 31050 1u4wp1y With Cover Page v2iswanNo ratings yet
- Kenalactams A–E from Nocardiopsis CG3 Show CytotoxicityDocument25 pagesKenalactams A–E from Nocardiopsis CG3 Show CytotoxicityOmar MessaoudiNo ratings yet
- Pharmacogenetic Determinants of Human Liver Microsomal Aminopyrine Metabolism and The Role of Cytochrome P450 2D6Document6 pagesPharmacogenetic Determinants of Human Liver Microsomal Aminopyrine Metabolism and The Role of Cytochrome P450 2D6Mediterr J Pharm Pharm SciNo ratings yet
- 1Document3 pages1Gina Manuela Torres AldanaNo ratings yet
- European Journal of Pharmaceutical Sciences: Mahmoud Kamel Mohamed, John Takyi-Williams, Bertrand Baudot, Anne GroblerDocument8 pagesEuropean Journal of Pharmaceutical Sciences: Mahmoud Kamel Mohamed, John Takyi-Williams, Bertrand Baudot, Anne GroblerLeidys Pérez SerpaNo ratings yet
- J. S. Fowler Et Al - ( 11-C) Spiroperidol: Synthesis, Specific Activity Determination, and Biodistribution in MiceDocument9 pagesJ. S. Fowler Et Al - ( 11-C) Spiroperidol: Synthesis, Specific Activity Determination, and Biodistribution in MiceLonkesNo ratings yet
- Chromatographic Separation of Piracetam and its MetaboliteDocument8 pagesChromatographic Separation of Piracetam and its MetaboliteFarhan J AhmadNo ratings yet
- Simon D. Brandt et al- Analytical chemistry of synthetic routes to psychoactive tryptamines Part 1: Characterisation of the Speeter and Anthony synthetic route to 5-methoxy-N,N-diisopropyltryptamine using ESI-MSMS and ESI-TOF-MSDocument11 pagesSimon D. Brandt et al- Analytical chemistry of synthetic routes to psychoactive tryptamines Part 1: Characterisation of the Speeter and Anthony synthetic route to 5-methoxy-N,N-diisopropyltryptamine using ESI-MSMS and ESI-TOF-MSJunmajNo ratings yet
- 38 FullDocument7 pages38 FullPaulomi PalNo ratings yet
- DOAs Tech and VickDocument4 pagesDOAs Tech and VickPrabhopNo ratings yet
- Huupponen 1992Document4 pagesHuupponen 1992aida aprianiNo ratings yet
- Radiosynthesis and biodistribution of dendrimer-avidin conjugate for cancer imagingDocument6 pagesRadiosynthesis and biodistribution of dendrimer-avidin conjugate for cancer imagingValentina RoznovNo ratings yet
- Analysis of Dithiocarbamate Fungicides in Vegetable Matrices Using HPLC-UV Followed by Atomic Absorption SpectrometryDocument7 pagesAnalysis of Dithiocarbamate Fungicides in Vegetable Matrices Using HPLC-UV Followed by Atomic Absorption SpectrometryaaNo ratings yet
- Jhet 58Document16 pagesJhet 58jason fiasonNo ratings yet
- Vitamin K3.2Document5 pagesVitamin K3.2lehuuhaiNo ratings yet
- Direct acetylation for full analysis of polysaccharides in edible plants andDocument12 pagesDirect acetylation for full analysis of polysaccharides in edible plants andmalvina.sekolonik911No ratings yet
- RCM 2100Document6 pagesRCM 2100Annia KurniawatiNo ratings yet
- Development of LC Method For Estimation of Diethyl Carbamazine Citrate and Chlorpheniramine Maleate in Combined Dosage FormDocument8 pagesDevelopment of LC Method For Estimation of Diethyl Carbamazine Citrate and Chlorpheniramine Maleate in Combined Dosage FormWhulan MudiaNo ratings yet
- Cuantificaciòn de CatinonasDocument8 pagesCuantificaciòn de CatinonasDFNo ratings yet
- HTTP WWW - Arkat-UsaDocument9 pagesHTTP WWW - Arkat-UsaGhayoor AbbasNo ratings yet
- Research PaperDocument8 pagesResearch PaperVeneta GizdakovaNo ratings yet
- 1-s2.0-S075333222201143X-mainDocument10 pages1-s2.0-S075333222201143X-mainhanNo ratings yet
- Comparative Single-Dose Pharmacokinetics of Amantadine Hydrochloride and Rimantadine Hydrochloride in Elderly AdultsDocument6 pagesComparative Single-Dose Pharmacokinetics of Amantadine Hydrochloride and Rimantadine Hydrochloride in Elderly AdultsLety LagunaNo ratings yet
- Journal of Chromatography B: Huaibing He, Ester Carballo-Jane, Xinchun Tong, Lucinda H. CohenDocument8 pagesJournal of Chromatography B: Huaibing He, Ester Carballo-Jane, Xinchun Tong, Lucinda H. CohenKuanNo ratings yet
- Helmy 2013Document5 pagesHelmy 2013adolfo olmosNo ratings yet
- Ge 2009Document3 pagesGe 2009RICHARD MACIENTE SILVINO DA SILVANo ratings yet
- Simultaneous Determination of Cefotaxime Sodium and Paracetamol by LC-MSDocument7 pagesSimultaneous Determination of Cefotaxime Sodium and Paracetamol by LC-MSIOSR Journal of PharmacyNo ratings yet
- Fukamachi Et Al-2008-Cancer ScienceDocument6 pagesFukamachi Et Al-2008-Cancer ScienceMonica Cabezas DipazNo ratings yet
- tmpB2A TMPDocument9 pagestmpB2A TMPFrontiersNo ratings yet
- Research ArticleDocument9 pagesResearch ArticleMinh Đức HoàngNo ratings yet
- Analytical Methods for Major and Modified Nucleosides - HPLC, GC, MS, NMR, UV and FT-IRFrom EverandAnalytical Methods for Major and Modified Nucleosides - HPLC, GC, MS, NMR, UV and FT-IRNo ratings yet
- Interglobe Technologies: About The CompanyDocument3 pagesInterglobe Technologies: About The Companyneeraj12121No ratings yet
- International Youth Day 2017 FlyerDocument1 pageInternational Youth Day 2017 Flyerneeraj12121No ratings yet
- A Glimpse of July 2018Document1 pageA Glimpse of July 2018neeraj12121No ratings yet
- DS18B20 (Temperature Sensor)Document20 pagesDS18B20 (Temperature Sensor)Kroos ZumNo ratings yet
- Y Division Nformation: EmissionDocument4 pagesY Division Nformation: Emissionneeraj12121No ratings yet
- Pinki Ke Joote PDFDocument28 pagesPinki Ke Joote PDFneeraj12121100% (1)
- Early Active Zone Assembly in Drosophila: DSyd-1 and DLiprin-α RoleDocument1 pageEarly Active Zone Assembly in Drosophila: DSyd-1 and DLiprin-α Roleneeraj12121No ratings yet
- Genus Statistics of The Virgo N-Body Simulations and The 1.2-Jy Redshift SurveyDocument20 pagesGenus Statistics of The Virgo N-Body Simulations and The 1.2-Jy Redshift Surveyneeraj12121No ratings yet
- DataDocument5 pagesDataneeraj12121No ratings yet
- CPC Cooperative Patent ClassificationDocument11 pagesCPC Cooperative Patent Classificationneeraj12121No ratings yet
- Artgan Content: Bout RojectDocument2 pagesArtgan Content: Bout Rojectneeraj12121No ratings yet
- IJCB2020 For Review TemplateDocument4 pagesIJCB2020 For Review Templateneeraj12121No ratings yet
- Reliable 4K Decoder Series with HDMI, VGA, BNC OutputsDocument8 pagesReliable 4K Decoder Series with HDMI, VGA, BNC Outputsneeraj12121No ratings yet
- Storage Choke, Fully Potted Resign: Description ApplicationsDocument3 pagesStorage Choke, Fully Potted Resign: Description Applicationsneeraj12121No ratings yet
- Chart Is Right For You? PDFDocument17 pagesChart Is Right For You? PDFميلاد نوروزي رهبرNo ratings yet
- Data Science Interview Questions: Answer HereDocument54 pagesData Science Interview Questions: Answer Hereneeraj12121No ratings yet
- Ray: A Distributed Framework For Emerging AI ApplicationsDocument19 pagesRay: A Distributed Framework For Emerging AI Applicationsneeraj12121No ratings yet
- Human Activity RecoDocument17 pagesHuman Activity Reconeeraj12121No ratings yet
- SVM PresentationDocument27 pagesSVM Presentationneeraj12121No ratings yet
- Tune: A Research Platform For Distributed Model Selection and TrainingDocument8 pagesTune: A Research Platform For Distributed Model Selection and Trainingneeraj12121No ratings yet
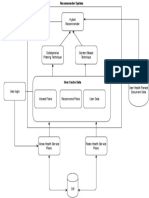
- Functional Flow PDFDocument1 pageFunctional Flow PDFneeraj12121No ratings yet
- Machine Learning Model Optimization with Gradient BoostingDocument17 pagesMachine Learning Model Optimization with Gradient Boostingneeraj12121No ratings yet
- Gradient BoostingDocument80 pagesGradient BoostingV Prasanna ShrinivasNo ratings yet
- Principal Component Analysis (PCA) : Dimensionality Reduction Using PcaDocument21 pagesPrincipal Component Analysis (PCA) : Dimensionality Reduction Using Pcaneeraj12121No ratings yet
- Hypertension: Key FactsDocument4 pagesHypertension: Key FactsGhaniNo ratings yet
- Shine-On 22Document3 pagesShine-On 22I Gusti PradanaNo ratings yet
- Cleft Lip and Palate: Protocol For Orthodontics Treatment: October 2021Document13 pagesCleft Lip and Palate: Protocol For Orthodontics Treatment: October 2021Aditya RamadhanNo ratings yet
- Nistami RTD SlidesDocument59 pagesNistami RTD Slidesrajantehri143No ratings yet
- YDEW3004-Case Study-316103481Document15 pagesYDEW3004-Case Study-316103481Jenelle Lee ColeNo ratings yet
- Fosroc Trafficguard UR200: Constructive SolutionsDocument2 pagesFosroc Trafficguard UR200: Constructive SolutionsVincent JavateNo ratings yet
- Rapid-p0014-Oui-Apc-ltr-0230 Ouijv Rigging and Lifting Procedure RapidDocument153 pagesRapid-p0014-Oui-Apc-ltr-0230 Ouijv Rigging and Lifting Procedure RapidYo Wee LiamNo ratings yet
- Unit 2 - Develop and Implement The Health and Safety Policy Workbook RGDocument11 pagesUnit 2 - Develop and Implement The Health and Safety Policy Workbook RGAshraf EL WardajiNo ratings yet
- Studi Kualitatif: Pelaksanaan Program Dukungan Psikososial Oleh Relawan Pada Bencana Gempa Bumi Di Lombok, Nusa Tenggara BaratDocument9 pagesStudi Kualitatif: Pelaksanaan Program Dukungan Psikososial Oleh Relawan Pada Bencana Gempa Bumi Di Lombok, Nusa Tenggara Barattaken takenbywclNo ratings yet
- Indonesia Healthcare ReportDocument51 pagesIndonesia Healthcare ReportPutri CandraNo ratings yet
- Regional Family Planning Inventory FormsDocument2 pagesRegional Family Planning Inventory FormsRANDY BAOGBOGNo ratings yet
- Proklenz One - MSDSDocument2 pagesProklenz One - MSDSneoNo ratings yet
- Perspectives of IDD in AsiaDocument7 pagesPerspectives of IDD in AsiaKura ChingNo ratings yet
- Bonnie Patten Article - Self-RegulationDocument24 pagesBonnie Patten Article - Self-RegulationBill KeepNo ratings yet
- Nature of Conflict ExplainedDocument24 pagesNature of Conflict ExplainedJabrail AminNo ratings yet
- MDC COVID-19 Mandatory Health Safety Protocols Upon Work Resumption - V5 20200513Document13 pagesMDC COVID-19 Mandatory Health Safety Protocols Upon Work Resumption - V5 20200513Clarabelle Mae Dela RosaNo ratings yet
- Hyland's 4 Kids Day & Night Cold 'N Cough Relief Liquid - 8 FL Oz TargetDocument1 pageHyland's 4 Kids Day & Night Cold 'N Cough Relief Liquid - 8 FL Oz TargetKarma JigmeyNo ratings yet
- Test Bank For Health Psychology 2 e 2nd Edition Deborah Fish RaginDocument34 pagesTest Bank For Health Psychology 2 e 2nd Edition Deborah Fish RaginpudgyoverburnnggNo ratings yet
- Download Yatra App for Flight DetailsDocument3 pagesDownload Yatra App for Flight DetailsShafi Moznur AhmedNo ratings yet
- Karen Horney Theorist in Psychoanalysis and Feminine PsychlogyDocument14 pagesKaren Horney Theorist in Psychoanalysis and Feminine PsychlogyaelindahoNo ratings yet
- 167 Manuscript 193 1 10 20210107Document4 pages167 Manuscript 193 1 10 20210107Hundessa Daba100% (1)
- Edgar Allan Poe - The Facts in The Case of M. ValdemarDocument13 pagesEdgar Allan Poe - The Facts in The Case of M. ValdemarCem AvciNo ratings yet
- LifeextensionDocument84 pagesLifeextensionhermestriNo ratings yet
- Guide Risk Assessment SMEsDocument29 pagesGuide Risk Assessment SMEsXozanNo ratings yet
- Strategy Implementation in Organizations A ConceptDocument14 pagesStrategy Implementation in Organizations A ConceptGemechis LemaNo ratings yet
- SDS-Dolphin Screen WashDocument10 pagesSDS-Dolphin Screen WashLiz CNo ratings yet
- Public Workshop #1 PresentationDocument28 pagesPublic Workshop #1 PresentationEllyn SantiagoNo ratings yet