You might also like
- ACLS Post TestDocument6 pagesACLS Post TestSheila Tolentino-Belanio73% (78)
- Elizabeth G. Querubin BSN 3E1-9 - Group 195 A Ectopic PregnancyDocument16 pagesElizabeth G. Querubin BSN 3E1-9 - Group 195 A Ectopic PregnancyLizeth Querubin97% (38)
- HSB Mock Paper 2Document17 pagesHSB Mock Paper 2Nadege Roach50% (2)
- Cognitive Function, Physical Activity, and Aging: Possible Biological Links and Implications For Multimodal InterventionsDocument17 pagesCognitive Function, Physical Activity, and Aging: Possible Biological Links and Implications For Multimodal Interventionskatherine monsalveNo ratings yet
- Appi Focus 20160030Document9 pagesAppi Focus 20160030Ners EducationNo ratings yet
- Experience During Adolescence Shapes Brain Development 2019Document20 pagesExperience During Adolescence Shapes Brain Development 2019adrianaNo ratings yet
- Neuropsychological Rehabilitation For Traumatic Brain Injury Patients PDFDocument12 pagesNeuropsychological Rehabilitation For Traumatic Brain Injury Patients PDFFausto D. GomezNo ratings yet
- SI Altera Comportamiento-Eje Mic-Circuitos NeuroquimicosDocument21 pagesSI Altera Comportamiento-Eje Mic-Circuitos NeuroquimicosCRISTOFER EDUARDO GONZALEZ IRARRAZABALNo ratings yet
- Defining Cognitive Reserve and Implications For Cognitive AgingDocument12 pagesDefining Cognitive Reserve and Implications For Cognitive AgingmarleimanNo ratings yet
- Mecanismos de Flexibilidad Cognitiva en Adolescentes Con TEADocument45 pagesMecanismos de Flexibilidad Cognitiva en Adolescentes Con TEAPsicoterapia InfantilNo ratings yet
- Cognitive ElementsDocument7 pagesCognitive Elementssolidus400No ratings yet
- Immune Modulation of Learning, Memory, Neural Plasticity and NeurogenesisDocument33 pagesImmune Modulation of Learning, Memory, Neural Plasticity and NeurogenesisRiuin23No ratings yet
- A Neurocognitive Approach To Understanding The Neurobiology of AddictionDocument7 pagesA Neurocognitive Approach To Understanding The Neurobiology of AddictionalejandraNo ratings yet
- Marquez-Chin-Popovic2020 Article FunctionalElectricalStimulatioDocument25 pagesMarquez-Chin-Popovic2020 Article FunctionalElectricalStimulatioMohamed Ali ZorkotNo ratings yet
- Irwin Slavich Psychoneuroimmunology 2017Document23 pagesIrwin Slavich Psychoneuroimmunology 2017meriem boualemNo ratings yet
- The Basics of Brain Development: ReviewDocument22 pagesThe Basics of Brain Development: ReviewmikimunNo ratings yet
- Aging and Immunity - Impact of Behavioral InterventionDocument15 pagesAging and Immunity - Impact of Behavioral InterventionAnnisaastikaNo ratings yet
- Van Den Bergh 2011 Li 2003: Anatomical MaturationDocument12 pagesVan Den Bergh 2011 Li 2003: Anatomical MaturationTitin Tria UtamiNo ratings yet
- Stern 2019Document6 pagesStern 2019marleimanNo ratings yet
- Pascual-Leone - Plasticity Across Lifespan - BTOP 2011Document14 pagesPascual-Leone - Plasticity Across Lifespan - BTOP 2011Carla Guixé SendiuNo ratings yet
- Garcia Garcia Et Al BrainhealthDocument26 pagesGarcia Garcia Et Al BrainhealthTheawakenedmindsNo ratings yet
- Health Psychology 10th Edition Taylor Solution ManualDocument7 pagesHealth Psychology 10th Edition Taylor Solution Manualcarlo100% (25)
- Environmental Influence in The Brain, Human Welfare and Mental HealthDocument11 pagesEnvironmental Influence in The Brain, Human Welfare and Mental HealthericjimenezNo ratings yet
- Epigenetics and Psychoneuroinmunology. Mechanims and ModelsDocument15 pagesEpigenetics and Psychoneuroinmunology. Mechanims and ModelsSparkniano HeNo ratings yet
- Aging Review RecomendadoDocument22 pagesAging Review RecomendadoseapeleoNo ratings yet
- Brainsci 13 01610 With CoverDocument33 pagesBrainsci 13 01610 With Coverbianki.treeNo ratings yet
- Ijbsv 19 P 3383Document12 pagesIjbsv 19 P 3383Sumeet TripathiNo ratings yet
- JNR 24151Document16 pagesJNR 24151Vinnie Juliana YonatanNo ratings yet
- Neuroimage: ClinicalDocument13 pagesNeuroimage: ClinicalJuli DeviaNo ratings yet
- 2010 - Neural Mechanisms of Ageing and Cognitive DeclineDocument19 pages2010 - Neural Mechanisms of Ageing and Cognitive DeclineВладимир ДружининNo ratings yet
- Efeitos Combinados Do Exercicio Dieta e Sono Sobre A Neuroplasticidade - 2022Document17 pagesEfeitos Combinados Do Exercicio Dieta e Sono Sobre A Neuroplasticidade - 2022Psicólogo Luis BernardoNo ratings yet
- The Role of Epigenetics in Psychological ResilienceDocument10 pagesThe Role of Epigenetics in Psychological ResilienceFrancisco MtzNo ratings yet
- Unbalanced Neuronal Circuits in AddictionDocument10 pagesUnbalanced Neuronal Circuits in AddictionWalter VillalobosNo ratings yet
- Enhancing Intelligence: From The Group To The IndividualDocument20 pagesEnhancing Intelligence: From The Group To The IndividualSebastiánsillo SánchezNo ratings yet
- Noack 09bDocument20 pagesNoack 09bSofia AreizaNo ratings yet
- Hill (2004) - Executive Dysfunction in AutismDocument7 pagesHill (2004) - Executive Dysfunction in Autismgabriela toroNo ratings yet
- Epigenetic SocialDocument5 pagesEpigenetic SocialClaudiu HbkNo ratings yet
- Davidson Social NNDocument7 pagesDavidson Social NNmagalyflorNo ratings yet
- Module 1 - NeuropsychologyDocument4 pagesModule 1 - Neuropsychologyaysha farihaNo ratings yet
- Post Op Cognitive DysfunctionDocument23 pagesPost Op Cognitive Dysfunctionlakshminivas PingaliNo ratings yet
- Social Circuits and Their Dysfunction in Autism Spectrum DisorderDocument13 pagesSocial Circuits and Their Dysfunction in Autism Spectrum DisorderMerry BearrNo ratings yet
- 1 s2.0 S088915910700027X Main PDFDocument10 pages1 s2.0 S088915910700027X Main PDFnermal93No ratings yet
- Review: Predicting Outcome After Childhood Brain InjuryDocument8 pagesReview: Predicting Outcome After Childhood Brain InjuryLuis Andrade ParraNo ratings yet
- 2023 The Role of Exerkines On Brain Mitochondria A Mini-ReviewDocument8 pages2023 The Role of Exerkines On Brain Mitochondria A Mini-Revieworellanaroco.10No ratings yet
- Grand Ner 2017Document22 pagesGrand Ner 2017EtaursaeNo ratings yet
- Human Social GenomicsDocument7 pagesHuman Social Genomicsjuan orjuelaNo ratings yet
- No. 30 Dafpus PDFDocument7 pagesNo. 30 Dafpus PDFItsAHZ TimeNo ratings yet
- Papel de La Microglía en La SinapsisDocument13 pagesPapel de La Microglía en La SinapsisLorena de la FlorNo ratings yet
- Cambios Relacionados Con La Edad en La Salud Cerebrovascular y Sus Efectos Sobre La Función Neuronal y La CogniciónDocument40 pagesCambios Relacionados Con La Edad en La Salud Cerebrovascular y Sus Efectos Sobre La Función Neuronal y La CogniciónDjc34No ratings yet
- 3 Beyond Origins: Developmental Pathways and The Dynamics of Brain NetworksDocument36 pages3 Beyond Origins: Developmental Pathways and The Dynamics of Brain NetworksSebastian PereiraNo ratings yet
- Bonetto 2021 Myelin - AgatekeeperofactivityDocument16 pagesBonetto 2021 Myelin - AgatekeeperofactivityMary KarabkNo ratings yet
- Rehabilitation For Stroke Behavioral, Temporal, and Spatial Targets For Cellular Transplants As Adjuncts ToDocument9 pagesRehabilitation For Stroke Behavioral, Temporal, and Spatial Targets For Cellular Transplants As Adjuncts ToCobzaru AlexandruNo ratings yet
- NeurotransmitterSchizophrenia KJS Indranath v2Document28 pagesNeurotransmitterSchizophrenia KJS Indranath v2eva pravitasari nefertitiNo ratings yet
- 14growth, Development and AgingDocument16 pages14growth, Development and AgingLailaYaned Gonzalez BejaranoNo ratings yet
- Characterizing - Cognitive - Aging - of - Working - Memory - 2014 - Córtex Pre FrontalDocument15 pagesCharacterizing - Cognitive - Aging - of - Working - Memory - 2014 - Córtex Pre FrontalDianna SantosNo ratings yet
- Synaptic Plasticity in Alzheimer's Disease and Healthy AgingDocument24 pagesSynaptic Plasticity in Alzheimer's Disease and Healthy AgingPAULA ITZEL AVALOS POLANCONo ratings yet
- 1 s2.0 S0006322315001626 MainDocument12 pages1 s2.0 S0006322315001626 MainNeus Sangrós VidalNo ratings yet
- Neuroimage: Cory S. Inman, G. Andrew James, Stephan Hamann, Justin K. Rajendra, Giuseppe Pagnoni, Andrew J. ButlerDocument11 pagesNeuroimage: Cory S. Inman, G. Andrew James, Stephan Hamann, Justin K. Rajendra, Giuseppe Pagnoni, Andrew J. ButlerFrontiersNo ratings yet
- Complexity in Neuropsychological Assessments of Cognitive Impairment: A Network Analysis ApproachDocument12 pagesComplexity in Neuropsychological Assessments of Cognitive Impairment: A Network Analysis ApproachDiane MxNo ratings yet
- Mnemonic Similarity Task: A Tool For Assessing Hippocampal IntegrityDocument14 pagesMnemonic Similarity Task: A Tool For Assessing Hippocampal IntegrityEszter SzerencsiNo ratings yet
- Neurobiology of Social Behavior: Toward an Understanding of the Prosocial and Antisocial BrainFrom EverandNeurobiology of Social Behavior: Toward an Understanding of the Prosocial and Antisocial BrainNo ratings yet
- Reasoningchapter Ow Re Book PDFDocument21 pagesReasoningchapter Ow Re Book PDFucanNo ratings yet
- Mrazik 2010Document12 pagesMrazik 2010ucanNo ratings yet
- Colom 2002 Education WechslerFSIQandgDocument15 pagesColom 2002 Education WechslerFSIQandgucanNo ratings yet
- Gifted and Average-Ability Childrens Potential FoDocument15 pagesGifted and Average-Ability Childrens Potential FoucanNo ratings yet
- Intelligence and Its Relationship To Achievement: June 2015Document20 pagesIntelligence and Its Relationship To Achievement: June 2015ucanNo ratings yet
- Dutton 2014c PDFDocument8 pagesDutton 2014c PDFucanNo ratings yet
- Intelligence: Cashen M. Boccio, Kevin M. Beaver, Joseph A. SchwartzDocument8 pagesIntelligence: Cashen M. Boccio, Kevin M. Beaver, Joseph A. SchwartzucanNo ratings yet
- Commenting On The Great Debate General Abilities SDocument11 pagesCommenting On The Great Debate General Abilities SucanNo ratings yet
- Reasoningchapter Ow Re Book PDFDocument21 pagesReasoningchapter Ow Re Book PDFucanNo ratings yet
- Ability and Achievement Variables in Average Low ADocument17 pagesAbility and Achievement Variables in Average Low AucanNo ratings yet
- Measuring Reasoning Ability: January 2005Document21 pagesMeasuring Reasoning Ability: January 2005ucanNo ratings yet
- Understanding The Profoundly Gifted: November 2017Document18 pagesUnderstanding The Profoundly Gifted: November 2017ucanNo ratings yet
- Confirmatory Factor Structure of The Kaufman Asses PDFDocument30 pagesConfirmatory Factor Structure of The Kaufman Asses PDFucanNo ratings yet
- Intelligence Is What The Intelligence Test Measures. SeriouslyDocument5 pagesIntelligence Is What The Intelligence Test Measures. SeriouslyucanNo ratings yet
- Theory of Intelligence. New York: Allyn and BaconDocument1 pageTheory of Intelligence. New York: Allyn and BaconucanNo ratings yet
- Acute Myeloid Leukemia - AML: Clinical BackgroundDocument12 pagesAcute Myeloid Leukemia - AML: Clinical BackgroundNyxa AbdullaNo ratings yet
- Drug Study SitagliptinDocument2 pagesDrug Study SitagliptinAl Theó0% (1)
- NIAID HIV Language Guide - March 2020Document27 pagesNIAID HIV Language Guide - March 2020ithran khoNo ratings yet
- Map of Africa: Quick Facts About AfricaDocument11 pagesMap of Africa: Quick Facts About AfricaJoela CastilNo ratings yet
- DAFTAR PUSTAKA PU SAYANG Revisi 1Document5 pagesDAFTAR PUSTAKA PU SAYANG Revisi 1REGINANo ratings yet
- Disorders of Children and AdolescentsDocument13 pagesDisorders of Children and AdolescentsJay Mark Samson CabreraNo ratings yet
- Diabetes Mellitus A Medical History Journey (Public Health Practices, Methods and Policies) 1st Edition 2016 PDFDocument102 pagesDiabetes Mellitus A Medical History Journey (Public Health Practices, Methods and Policies) 1st Edition 2016 PDFsun sealNo ratings yet
- Gastric Case ClerkingDocument19 pagesGastric Case ClerkingAnelkaLouisNo ratings yet
- Reticuloendothelial SystemDocument26 pagesReticuloendothelial SystemNashreen QtqtNo ratings yet
- Antiarrhythmic Agent - WikipediaDocument23 pagesAntiarrhythmic Agent - WikipediaSai Jeevan SampathiraoNo ratings yet
- CPG PCP ListDocument5 pagesCPG PCP ListBryan AtasNo ratings yet
- Awareness of Fake Braces Usage Among Y-GenerationsDocument4 pagesAwareness of Fake Braces Usage Among Y-GenerationsdraNo ratings yet
- Seclusion and Physical Restraint: GeneralDocument6 pagesSeclusion and Physical Restraint: GeneralMegan ClearyNo ratings yet
- Orhan 2021 (Book) - Ultrasonography in Dentomaxillofacial Diagnostics - SpringerDocument372 pagesOrhan 2021 (Book) - Ultrasonography in Dentomaxillofacial Diagnostics - SpringerlagianggenNo ratings yet
- Icpc Heart ScreeningDocument2 pagesIcpc Heart ScreeningMahmoud TitilayoNo ratings yet
- Sankara Nethralaya Clinical Practice Patterns in OPHTHALMOLOGYDocument452 pagesSankara Nethralaya Clinical Practice Patterns in OPHTHALMOLOGYjordi28098380% (5)
- Laporan PBL ImunHema B Kelompok 10 Modul 2 Skenario 2Document32 pagesLaporan PBL ImunHema B Kelompok 10 Modul 2 Skenario 2Riswanto RuslanNo ratings yet
- 2021 Diabetes Clinical Practice GuidelineDocument15 pages2021 Diabetes Clinical Practice GuidelineAbdulfetah AhmedNo ratings yet
- PSM (Must Know)Document19 pagesPSM (Must Know)PranavNo ratings yet
- Playbook v3Document34 pagesPlaybook v3Rufaida MudrikaNo ratings yet
- Transfusion ReactionDocument6 pagesTransfusion ReactionHiraya ManawariNo ratings yet
- "My Neck Feels Stiff and There's Pain Coming From It. Every Time I Move It, It Only Gets Worse." As Stated by TheDocument4 pages"My Neck Feels Stiff and There's Pain Coming From It. Every Time I Move It, It Only Gets Worse." As Stated by TheCassey CuregNo ratings yet
- Nur 145 P1Document13 pagesNur 145 P1Kristine Mae LauritoNo ratings yet
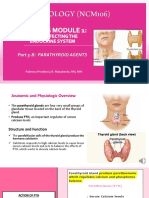
- Module 1 Part 5B. Parathyroid AgentsDocument21 pagesModule 1 Part 5B. Parathyroid AgentsBSN2G- SABLA-ON LORRAINE ANNENo ratings yet
- Symbicort Rapihaler (Budesonide and Formoterol Fumarate Dihydrate) Drug MediciDocument14 pagesSymbicort Rapihaler (Budesonide and Formoterol Fumarate Dihydrate) Drug Medicijr7mondo7edoNo ratings yet
- Introduction To Medical Microbiology Exam #1 Study GuideDocument1 pageIntroduction To Medical Microbiology Exam #1 Study GuideBangtan J-hopiaNo ratings yet
- Hypertension CASE REPORTDocument57 pagesHypertension CASE REPORTJulienne Sanchez-SalazarNo ratings yet