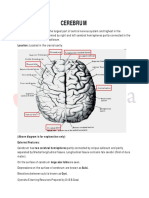
You might also like
- Module 1 - A Conversation About Reading and The Brain Study GuideDocument19 pagesModule 1 - A Conversation About Reading and The Brain Study Guideapi-530797247No ratings yet
- Convolutional Neural Networks in Python: Beginner's Guide to Convolutional Neural Networks in PythonFrom EverandConvolutional Neural Networks in Python: Beginner's Guide to Convolutional Neural Networks in PythonNo ratings yet
- Armaturi Profile PVCDocument53 pagesArmaturi Profile PVCstoian andreiNo ratings yet
- Anatomy and Physiology (Status Epilepticus)Document3 pagesAnatomy and Physiology (Status Epilepticus)Marvin John Labiano33% (3)
- Clinical Neuro-Ophthalmology-A Practical Guide-2007Document23 pagesClinical Neuro-Ophthalmology-A Practical Guide-2007joaquin mellaNo ratings yet
- Halaman DepanDocument1 pageHalaman DepanHesti hasanNo ratings yet
- Duchowski chpt2Document10 pagesDuchowski chpt2eimaio karxasNo ratings yet
- Gollisch MeisterDocument15 pagesGollisch MeisterElaheh LotfiNo ratings yet
- The RetinaDocument6 pagesThe RetinaSourav RanjanNo ratings yet
- Salazar - 2019 IntechOpen - Anatomy of The Human Optic Nerve - Structure and FuncionDocument46 pagesSalazar - 2019 IntechOpen - Anatomy of The Human Optic Nerve - Structure and FuncionLee Vo AnNo ratings yet
- ANATO - Campos VisuaisDocument11 pagesANATO - Campos VisuaishelbertpalmieroNo ratings yet
- Introduction To Neuropthalmology OptDocument25 pagesIntroduction To Neuropthalmology Opthenok birukNo ratings yet
- PhotopicDocument10 pagesPhotopicVivian AmosNo ratings yet
- Simple Anatomy of The RetinaDocument24 pagesSimple Anatomy of The RetinaLateepheart YakubNo ratings yet
- Visual Pathway 2021Document83 pagesVisual Pathway 2021Pavan chowdaryNo ratings yet
- Anatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterDocument20 pagesAnatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterAndres BenavidesNo ratings yet
- Anatomy of The Human Optic Nerve - Structure and FuncionDocument46 pagesAnatomy of The Human Optic Nerve - Structure and FuncionHesti hasanNo ratings yet
- Neuroanatomy, Pupillary Light Reflexes and PathwayDocument6 pagesNeuroanatomy, Pupillary Light Reflexes and PathwayMatthew PrestonNo ratings yet
- Apical Dendrite and Theory of Consiousness (La Berge, 2007)Document17 pagesApical Dendrite and Theory of Consiousness (La Berge, 2007)Gabriel ArteagaNo ratings yet
- For The Electronics Screen, See - For Other Uses, See .: Jump To Navigation Jump To SearchDocument10 pagesFor The Electronics Screen, See - For Other Uses, See .: Jump To Navigation Jump To SearchNicoleta TuleaNo ratings yet
- Photochemistry of VisionDocument35 pagesPhotochemistry of VisionHugh Jacobs60% (5)
- Perception NotesDocument12 pagesPerception NotesAnu KNo ratings yet
- Neurostiinte - Eye, Brain, Vision - David Hubel (Cap 3,4 p32-83) PDFDocument52 pagesNeurostiinte - Eye, Brain, Vision - David Hubel (Cap 3,4 p32-83) PDFgicutaNo ratings yet
- Chapter 3 - Uvea - 2012 - Clinical Anatomy and Physiology of The Visual SystemDocument21 pagesChapter 3 - Uvea - 2012 - Clinical Anatomy and Physiology of The Visual SystemAldo CortesNo ratings yet
- 2020OPTICNERVEPRESENTATIONDocument10 pages2020OPTICNERVEPRESENTATIONTendaiNo ratings yet
- Sreejith TDocument37 pagesSreejith TAbdisamed AllaaleNo ratings yet
- 1 Online PDFDocument18 pages1 Online PDFFera SherlianaNo ratings yet
- Macroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheDocument14 pagesMacroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheRoziel Anne BasaNo ratings yet
- Visual SystemDocument56 pagesVisual SystemMartha MulusaNo ratings yet
- A&p ActDocument4 pagesA&p ActGURPARABJOT KAURNo ratings yet
- Human Eye: A Human Eyeball Is Like A Simple Camera!Document18 pagesHuman Eye: A Human Eyeball Is Like A Simple Camera!ljieNo ratings yet
- Visual and Lacrimal AppratusDocument45 pagesVisual and Lacrimal AppratusAzman Bin KadirNo ratings yet
- Jurnal Anatomy of The Human Visual PathwayDocument7 pagesJurnal Anatomy of The Human Visual Pathwayhenry balanNo ratings yet
- BLOK 1.3 Jaras PenglihatanDocument43 pagesBLOK 1.3 Jaras PenglihatanlenypsNo ratings yet
- Basic AnotomyDocument48 pagesBasic AnotomyTadiwa MusaraNo ratings yet
- RETINADocument4 pagesRETINAElizabeth Anne Loto100% (1)
- Retina: I. CharacteristicsDocument4 pagesRetina: I. CharacteristicsElizabeth Anne LotoNo ratings yet
- Artificial Eye C 1 I: Hapter NtroductionDocument17 pagesArtificial Eye C 1 I: Hapter NtroductionswathichvNo ratings yet
- Artificial EyeDocument31 pagesArtificial Eyeapi-19937584No ratings yet
- The Physics of Light and Color - Human Vision and Color Perception - Olympus LSDocument14 pagesThe Physics of Light and Color - Human Vision and Color Perception - Olympus LSmarialopezmartinez424No ratings yet
- p88 99Document12 pagesp88 99Prasanna Venkhatesh VNo ratings yet
- Ocular and Visual Science: Nor Shahahidah BT Shahabudin Semester 3Document44 pagesOcular and Visual Science: Nor Shahahidah BT Shahabudin Semester 3sheedah_damuloNo ratings yet
- Introduction To The Visual System: Body, Which Produces The Components of The Aqueous Humor andDocument9 pagesIntroduction To The Visual System: Body, Which Produces The Components of The Aqueous Humor andUlziibayar BattaivanNo ratings yet
- Histology of Cornea & RetinaDocument14 pagesHistology of Cornea & RetinaSandhya KurupNo ratings yet
- Computer Vision: 03 - Neural - Operations - On - ImagesDocument21 pagesComputer Vision: 03 - Neural - Operations - On - ImagesDr. AnasNo ratings yet
- RD, M Hammam FaishalDocument29 pagesRD, M Hammam FaishalDesra AufarNo ratings yet
- Bio Psychology Lecture Notes Chapter 6 The Visual SystemDocument7 pagesBio Psychology Lecture Notes Chapter 6 The Visual SystemGeneric_Persona50% (2)
- Discovering The Wiring Diagram of The BrainDocument9 pagesDiscovering The Wiring Diagram of The Brainsiempreviva84No ratings yet
- The Anatomy of The LimbusDocument8 pagesThe Anatomy of The LimbusBukaNo ratings yet
- Anatomy of The Eye and OrbitDocument7 pagesAnatomy of The Eye and OrbitmmadomadNo ratings yet
- Orbit 2Document27 pagesOrbit 2cvmqx7yppdNo ratings yet
- Physiology Assignment - 085344Document10 pagesPhysiology Assignment - 085344Samuel A OlowolayemoNo ratings yet
- The EyeDocument36 pagesThe EyeMuresan Radu CatalinNo ratings yet
- Retina Anatomy - Overview, Gross Anatomy, Microscopic AnatomyDocument3 pagesRetina Anatomy - Overview, Gross Anatomy, Microscopic AnatomyjackNo ratings yet
- 21 Neural Control N Coordination-NCERT Questions N AnswersDocument6 pages21 Neural Control N Coordination-NCERT Questions N AnswersGuru SrinivaasNo ratings yet
- Specialized Sensory Organs: - EYE (Perceives Light)Document11 pagesSpecialized Sensory Organs: - EYE (Perceives Light)TodesengelNo ratings yet
- 1 - Newman1973Document16 pages1 - Newman1973Eduardo HernandezNo ratings yet
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Plates illustrating the natural and morbid changes of the human eyeFrom EverandPlates illustrating the natural and morbid changes of the human eyeNo ratings yet
- Convolutional Neural Networks: Fundamentals and Applications for Analyzing Visual ImageryFrom EverandConvolutional Neural Networks: Fundamentals and Applications for Analyzing Visual ImageryNo ratings yet
- Suprachoroidal Space InterventionsFrom EverandSuprachoroidal Space InterventionsShohista SaidkasimovaNo ratings yet
- Grave's Ophthalmopathy: Susy Fatmariyanti Delfitri Lutfi, Hendrian D. SoebagjoDocument12 pagesGrave's Ophthalmopathy: Susy Fatmariyanti Delfitri Lutfi, Hendrian D. SoebagjoriskhapangestikaNo ratings yet
- Laico: Ravind YE OspitalsDocument2 pagesLaico: Ravind YE OspitalsriskhapangestikaNo ratings yet
- Duty Report Friday, April 30, 2018: Ophthalmologist: Dr. Afrisal, SP.M (K) Senior: Dr. Soraya Junior: Dr. RiskhaDocument14 pagesDuty Report Friday, April 30, 2018: Ophthalmologist: Dr. Afrisal, SP.M (K) Senior: Dr. Soraya Junior: Dr. RiskhariskhapangestikaNo ratings yet
- Herpes Simplex KeratitisDocument20 pagesHerpes Simplex KeratitisriskhapangestikaNo ratings yet
- Pasien Jaga: 15 Desember 2018 Yet + DayDocument5 pagesPasien Jaga: 15 Desember 2018 Yet + DayriskhapangestikaNo ratings yet
- Eye Poster PromotionDocument1 pageEye Poster PromotionriskhapangestikaNo ratings yet
- Duty Report 30 Maret 2018Document43 pagesDuty Report 30 Maret 2018riskhapangestikaNo ratings yet
- Effective Cataract Surgical Coverage: An Indicator For Measuring Quality-Of-Care in The Context of Universal Health CoverageDocument13 pagesEffective Cataract Surgical Coverage: An Indicator For Measuring Quality-Of-Care in The Context of Universal Health CoverageriskhapangestikaNo ratings yet
- Efferent Visual System (Ocular Motor Pathways)Document38 pagesEfferent Visual System (Ocular Motor Pathways)riskhapangestika100% (1)
- Biochemistry of LensDocument24 pagesBiochemistry of LensriskhapangestikaNo ratings yet
- All Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-MullaDocument45 pagesAll Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-Mullariskhapangestika100% (1)
- Lobes of The Brain - FrontalDocument12 pagesLobes of The Brain - FrontalLoccos SienaNo ratings yet
- Design and Dosimetric Analysis of An Exposure Facility For Investigating Possible E¡ects of 2.45 GHzWi Fi Signals On Human SleepDocument11 pagesDesign and Dosimetric Analysis of An Exposure Facility For Investigating Possible E¡ects of 2.45 GHzWi Fi Signals On Human Sleepdarko6102No ratings yet
- Neuromythology of EINSTEIN BriainDocument5 pagesNeuromythology of EINSTEIN BriainrorozainosNo ratings yet
- The Central Nervous System: Chapter 12 Part ADocument31 pagesThe Central Nervous System: Chapter 12 Part AMajdi AkNo ratings yet
- The Inter-Relationships Between Cerebral Visual Impairment, Autism and Intellectual DisabilityDocument10 pagesThe Inter-Relationships Between Cerebral Visual Impairment, Autism and Intellectual DisabilityShobithaNo ratings yet
- 15 CerebrumDocument32 pages15 CerebrumMédecin Adrian TGNo ratings yet
- NetVR Version 1.3 Supported Cameras and EncodersDocument369 pagesNetVR Version 1.3 Supported Cameras and EncodersBraulio Andres Arenas VillavicencioNo ratings yet
- Benton Visual Retention TestDocument53 pagesBenton Visual Retention TestRin Sua100% (1)
- Labores Interior MinaDocument75 pagesLabores Interior MinaFernandoNo ratings yet
- HAMR ManualDocument120 pagesHAMR Manualwarclone100% (3)
- Patient EVRDocument12 pagesPatient EVRJorge CastilloNo ratings yet
- Altitudinal Visual Field Defect (Bombay)Document2 pagesAltitudinal Visual Field Defect (Bombay)Moileon2011No ratings yet
- CatalogMobilier Urban 2Document172 pagesCatalogMobilier Urban 2StefiLupanNo ratings yet
- Cerebrovascular AccidentDocument53 pagesCerebrovascular AccidentJulia Salvio50% (2)
- 19 FN - GL, XNDocument310 pages19 FN - GL, XNAnonymous Tvfioo9MQ8No ratings yet
- Ebook Psychology Psyc2020 Psyc2021 Custom Edition by Schutte E Ed Et A PDF Full Chapter PDFDocument67 pagesEbook Psychology Psyc2020 Psyc2021 Custom Edition by Schutte E Ed Et A PDF Full Chapter PDFedward.bittner820100% (33)
- Cerebral Mechanisms of Hypnotic Induction and SuggestionDocument29 pagesCerebral Mechanisms of Hypnotic Induction and SuggestionArnt van HeldenNo ratings yet
- Visual Pathway & Its LesionsDocument24 pagesVisual Pathway & Its LesionsSyedaNaveenBatoolNo ratings yet
- Perceptual Pleasure and The Brain by Irving Biederman & Edward VesselDocument8 pagesPerceptual Pleasure and The Brain by Irving Biederman & Edward VesselAmiraNo ratings yet
- Sway TrumpetDocument1 pageSway TrumpetOleg ZaiatNo ratings yet
- Virtual Reality For Neurorehabilitation and Cognitive EnhancementDocument20 pagesVirtual Reality For Neurorehabilitation and Cognitive EnhancementAndrés RodríguezNo ratings yet
- Lecture - Slides-01 - 06 - Medial Surface of The BrainDocument6 pagesLecture - Slides-01 - 06 - Medial Surface of The BrainMaria Eduarda VianaNo ratings yet
- Cerebrum 130906013333Document12 pagesCerebrum 130906013333RajveerNo ratings yet
- Tracker December 01 To 12TH 2020Document12 pagesTracker December 01 To 12TH 2020Nasir AhmedNo ratings yet
- Dissection of The Sheep'S BrainDocument13 pagesDissection of The Sheep'S BrainBivolaru AlinNo ratings yet
- 5 Optic RadiationsDocument2 pages5 Optic RadiationsJennifer ConradNo ratings yet
- Assignment 3 Anurag Satpathy 116EE0343: For 3 IterationsDocument5 pagesAssignment 3 Anurag Satpathy 116EE0343: For 3 IterationsAnurag SatpathyNo ratings yet