You might also like
- Pulse and Fourier Transform NMR: Introduction to Theory and MethodsFrom EverandPulse and Fourier Transform NMR: Introduction to Theory and MethodsNo ratings yet
- Aliment at Ore Alinco - DM330MVE - SMDocument14 pagesAliment at Ore Alinco - DM330MVE - SMdaniele_mdgNo ratings yet
- Trower, 1996. in Vitro Mutagenesis Protocols Volume 57Document9 pagesTrower, 1996. in Vitro Mutagenesis Protocols Volume 57afinaNo ratings yet
- Keiji Nakajima Et Al - Opposite Stereospecificity of Two Tropinone Reductases Is Conferred by The Substrate-Binding SiteDocument4 pagesKeiji Nakajima Et Al - Opposite Stereospecificity of Two Tropinone Reductases Is Conferred by The Substrate-Binding SiteHumiopNo ratings yet
- DNA/RNA Real-Time Quantitative PCRDocument8 pagesDNA/RNA Real-Time Quantitative PCRbiosynthesis12No ratings yet
- (Medika) Real-Time PCR System-Gentier CatalogDocument78 pages(Medika) Real-Time PCR System-Gentier CatalogRidayat SisNo ratings yet
- Capabilities and Limitations of Multirate Control SchemesDocument11 pagesCapabilities and Limitations of Multirate Control SchemesSWATI SINGHNo ratings yet
- (ASN) Real-Time PCR System-Gentier Catalog-1Document78 pages(ASN) Real-Time PCR System-Gentier Catalog-1Ridayat SisNo ratings yet
- Form KerohanianDocument2 pagesForm KerohanianAlfian AgustamaNo ratings yet
- PCR - Real TimeDocument21 pagesPCR - Real TimeChua KhachychyNo ratings yet
- Cu41277 20210910 0001Document6 pagesCu41277 20210910 0001Abdullah al ImranNo ratings yet
- 3 Okt 2019Document34 pages3 Okt 2019ElitaNo ratings yet
- Adobe Scan 11 Mar 2023Document12 pagesAdobe Scan 11 Mar 2023Indra “The starter” OtsutsukiNo ratings yet
- Es Lab Manual Next Exp.Document11 pagesEs Lab Manual Next Exp.AnmolGoyalNo ratings yet
- 2022-04-26 L5 - Annealing Temperature & Primer DesignDocument3 pages2022-04-26 L5 - Annealing Temperature & Primer DesignTamara ElyasNo ratings yet
- JCM 32 4 1095-1098 1994Document4 pagesJCM 32 4 1095-1098 1994mariotecNo ratings yet
- Transcription: The Path and Control of Gene ExpressionDocument24 pagesTranscription: The Path and Control of Gene ExpressionPaolo NaguitNo ratings yet
- Perrey 1998Document5 pagesPerrey 1998Shukr Wesman BlbasNo ratings yet
- 1994 - Jones, Franklin, Thomas - Molecular analysis of the operon which encodes the RNA polymerase sigma factor σ54 of Escherichia coliDocument9 pages1994 - Jones, Franklin, Thomas - Molecular analysis of the operon which encodes the RNA polymerase sigma factor σ54 of Escherichia coliPJ Chalet BookNo ratings yet
- R831 Air Water Heat PumpDocument62 pagesR831 Air Water Heat PumpRodrigoCisnerosNo ratings yet
- Chapter 5b Molecular Genetics Part 2Document63 pagesChapter 5b Molecular Genetics Part 2biñan lagunaNo ratings yet
- Differential DisplayDocument25 pagesDifferential DisplayElsayed Refaat Aly MareyNo ratings yet
- Polymerase Chain Reaction (PCR)Document17 pagesPolymerase Chain Reaction (PCR)pushpavalliNo ratings yet
- LiIO3 LiNbO3Document5 pagesLiIO3 LiNbO3javiervrNo ratings yet
- Haruyama1986-Thin-Film Platinum Resistance Thermometer For Use at Low Temperatures and in High Magnetic FieldsDocument3 pagesHaruyama1986-Thin-Film Platinum Resistance Thermometer For Use at Low Temperatures and in High Magnetic FieldsAlexandre HenriquesNo ratings yet
- TranscriptionDocument19 pagesTranscriptionNAVYA A/P SOORIA THEYMUDUNo ratings yet
- The Key Steps of TranscriptionDocument13 pagesThe Key Steps of TranscriptionbiochemNo ratings yet
- McrA Expression GeneDocument7 pagesMcrA Expression GenelaksiyerNo ratings yet
- Mueller Military DocumentsDocument16 pagesMueller Military DocumentsDan LamotheNo ratings yet
- NRL Report Analyzes Experimental Crossed-Loop Antennas for Submarine VLF ReceptionDocument40 pagesNRL Report Analyzes Experimental Crossed-Loop Antennas for Submarine VLF ReceptioncsezenogluNo ratings yet
- Catalogo General Alco RSD 16Document355 pagesCatalogo General Alco RSD 16Ivan100% (1)
- Speed 13Document1 pageSpeed 13docpot2008No ratings yet
- 619e4d83b653b Biology Genetics Laboratory Exercise The Central Dogma of GeneticsDocument10 pages619e4d83b653b Biology Genetics Laboratory Exercise The Central Dogma of GeneticsKimNo ratings yet
- Et Al., Et Al., Et Al., Et Ai., Et Al., Et Al., Et Al.Document7 pagesEt Al., Et Al., Et Al., Et Ai., Et Al., Et Al., Et Al.Walid BadrNo ratings yet
- Jolivet Concertino - Trumpet PartDocument7 pagesJolivet Concertino - Trumpet Partedison samuel100% (1)
- TRW - Mutasi Dan DNA RepairDocument49 pagesTRW - Mutasi Dan DNA RepairYUFFANo ratings yet
- Subunidad SigmaDocument5 pagesSubunidad SigmaGeorgina HernandezNo ratings yet
- Brosur HbA1c & CRP DxGENDocument1 pageBrosur HbA1c & CRP DxGENRini HarwatiNo ratings yet
- Rna Interference: PCR Strategies For The Quantification of Stable Degradation-Fragments Derived From Sirna-Targeted MrnasDocument5 pagesRna Interference: PCR Strategies For The Quantification of Stable Degradation-Fragments Derived From Sirna-Targeted Mrnasbiotecno1No ratings yet
- IAS-Botany-Paper-2-2020Document6 pagesIAS-Botany-Paper-2-2020kanchanbirat100No ratings yet
- Nar 00609 H 2013 File006Document9 pagesNar 00609 H 2013 File006Ronilo Jose Danila FloresNo ratings yet
- Medical HistoryDocument7 pagesMedical HistorySuryAdi PianoNo ratings yet
- Real Time PCR: Pramod Bahadur K.CDocument14 pagesReal Time PCR: Pramod Bahadur K.CPramod KcNo ratings yet
- cross talk measuringDocument8 pagescross talk measuringprashant PujariNo ratings yet
- Speed 08Document1 pageSpeed 08docpot2008No ratings yet
- Cbse 2009 Physics Board Paper Class XIIDocument17 pagesCbse 2009 Physics Board Paper Class XIINeil Mahaseth86% (7)
- Aplikasi Teknik Biologi Molekuler - CompressedDocument31 pagesAplikasi Teknik Biologi Molekuler - CompressedHelmawati PasombaNo ratings yet
- Normal ValuesDocument6 pagesNormal ValuesVenkat TNo ratings yet
- Central Dogma Review KEYDocument8 pagesCentral Dogma Review KEYeula faith miracle andam0% (1)
- Century Packers Door to Door Cargo Shipping RatesDocument1 pageCentury Packers Door to Door Cargo Shipping Ratesrocky romeoNo ratings yet
- DNA transcription - Regulation of gene expression through RNA polymeraseDocument38 pagesDNA transcription - Regulation of gene expression through RNA polymerasetoobashafiNo ratings yet
- BC5051 PCR Lecture EKDocument24 pagesBC5051 PCR Lecture EKLavinia MihaiNo ratings yet
- ACFrOgDWGR7iXyIO5GsdkRtH3Exx5VlMTm1ltALd8ms37CeVjv9YaiNPOHzcK0nUC FX8qH3sSuYeYEltAe KuPE1xyq6MOkpGx9ZKKbFui3xpyDepIk93 KGP VRsZIHpQNt8p4vnTrgvUzujsTDocument9 pagesACFrOgDWGR7iXyIO5GsdkRtH3Exx5VlMTm1ltALd8ms37CeVjv9YaiNPOHzcK0nUC FX8qH3sSuYeYEltAe KuPE1xyq6MOkpGx9ZKKbFui3xpyDepIk93 KGP VRsZIHpQNt8p4vnTrgvUzujsTAniket AmanNo ratings yet
- Antologia 2-3 ViolinDocument46 pagesAntologia 2-3 Violinanmari89No ratings yet
- 8310 01 VHF2048Document93 pages8310 01 VHF2048Claudemir VianaNo ratings yet
- A Novel Method For Real Time Quantitative RT-PCRDocument8 pagesA Novel Method For Real Time Quantitative RT-PCRSkidi pap papNo ratings yet
- Dalma Z 1976Document16 pagesDalma Z 1976Redouane idrissiNo ratings yet
- Adobe Scan 12 Feb 2024Document1 pageAdobe Scan 12 Feb 2024Rohit BansalNo ratings yet
- Nha DrawingDocument23 pagesNha DrawingEngr Nissar KakarNo ratings yet
- PCX - Uer I: Ampl Fy S DevDocument55 pagesPCX - Uer I: Ampl Fy S DevJagadeeshwaran MNo ratings yet
- Protein Synthesis NotesDocument6 pagesProtein Synthesis NotesRokunuz Jahan RudroNo ratings yet
- RNA Polymerases and Eukaryotic TranscriptionDocument5 pagesRNA Polymerases and Eukaryotic TranscriptionTadhg Ó MaoldhomhnaighNo ratings yet
- Basic Molecular Genetic Mechanisms: (Part 3)Document35 pagesBasic Molecular Genetic Mechanisms: (Part 3)ofirNo ratings yet
- Nuclei C Acids: BSN 1y0-5Document25 pagesNuclei C Acids: BSN 1y0-5joevette_30No ratings yet
- VDJ Recombination With Antibody StructureDocument38 pagesVDJ Recombination With Antibody Structuresuperfr3shmNo ratings yet
- Explore How Gene Mutations Alter Protein SynthesisDocument4 pagesExplore How Gene Mutations Alter Protein SynthesisSamuel Morales NavarroNo ratings yet
- Molecular diversity analysis of wheat genotypes using RAPD and SSR markersDocument8 pagesMolecular diversity analysis of wheat genotypes using RAPD and SSR markersHarshvardhan ZalaNo ratings yet
- Clicker QuestionsDocument29 pagesClicker QuestionsSally LeeNo ratings yet
- Nextgen ExplanationDocument14 pagesNextgen ExplanationecogandhiNo ratings yet
- Transcription ProcessDocument85 pagesTranscription Processshruti shahNo ratings yet
- Definition of A Human Being HandoutDocument11 pagesDefinition of A Human Being HandoutSally Jo Sorensen100% (4)
- DNA and RNA StructureDocument4 pagesDNA and RNA StructureH DNo ratings yet
- Dna Repair MechanismsDocument49 pagesDna Repair MechanismsPurushottam GuptaNo ratings yet
- DNA Replication and RepairDocument7 pagesDNA Replication and RepairJhun Lerry TayanNo ratings yet
- Maxwell Forensic Labs Brochure Promega PDFDocument8 pagesMaxwell Forensic Labs Brochure Promega PDFAhmad HamdounNo ratings yet
- Diseño de Oligos Hector NeriDocument3 pagesDiseño de Oligos Hector NeriHECTOR DANIEL NERI MAGALLANESNo ratings yet
- Philippine high school pre-final exam on science, hormones, DNA, RNA and protein synthesisDocument2 pagesPhilippine high school pre-final exam on science, hormones, DNA, RNA and protein synthesisRaquel Bona ViñasNo ratings yet
- UST Bio Exam Part ReviewerDocument6 pagesUST Bio Exam Part ReviewerAmanda PoetirayNo ratings yet
- Essay Question Protein SynthesisDocument8 pagesEssay Question Protein Synthesisalexismillerspringfield100% (2)
- Paper I Unit Ii:-Gene Expression Rna Polymerase DefinitionDocument53 pagesPaper I Unit Ii:-Gene Expression Rna Polymerase DefinitionRithwik ZNo ratings yet
- Samarpan BiologyDocument23 pagesSamarpan Biologyblack kobraNo ratings yet
- Chenqi Liu - 11 - DNA Vs RNA and Protein Synthesis - Amoeba Sisters - SHODocument2 pagesChenqi Liu - 11 - DNA Vs RNA and Protein Synthesis - Amoeba Sisters - SHOthomasmarteauhNo ratings yet
- tRNA-derived Small RNAs Target Transposable Element TranscriptsDocument11 pagestRNA-derived Small RNAs Target Transposable Element TranscriptsMan Ba SotzNo ratings yet
- Epigenetics - A New Frontier in DentistryDocument11 pagesEpigenetics - A New Frontier in DentistryDiego Andres Hincapie HerreraNo ratings yet
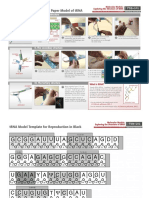
- Trna Model PDFDocument3 pagesTrna Model PDFYoga NovalNo ratings yet
- Difference Between Dna & Rna: Students NameDocument5 pagesDifference Between Dna & Rna: Students NameHisham MustafaNo ratings yet
- Unit 8 - Week 5: Assignment 5Document4 pagesUnit 8 - Week 5: Assignment 5Ananda VijayasarathyNo ratings yet
- The E. Coli Insulin FactoryDocument7 pagesThe E. Coli Insulin FactoryKEZIAH HERRERA0% (1)
- Transcription and TranslationDocument35 pagesTranscription and TranslationMing mingNo ratings yet
- TranslationDocument37 pagesTranslationFarhana AnuarNo ratings yet
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolFrom EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolNo ratings yet
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsFrom EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsRating: 4 out of 5 stars4/5 (146)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeFrom EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (3)
- Meltdown: Nuclear disaster and the human cost of going criticalFrom EverandMeltdown: Nuclear disaster and the human cost of going criticalRating: 5 out of 5 stars5/5 (5)
- Guidelines for Asset Integrity ManagementFrom EverandGuidelines for Asset Integrity ManagementRating: 5 out of 5 stars5/5 (1)
- Coating and Drying Defects: Troubleshooting Operating ProblemsFrom EverandCoating and Drying Defects: Troubleshooting Operating ProblemsRating: 5 out of 5 stars5/5 (1)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilFrom EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilRating: 5 out of 5 stars5/5 (1)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeFrom EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeRating: 5 out of 5 stars5/5 (1)
- Gas-Liquid And Liquid-Liquid SeparatorsFrom EverandGas-Liquid And Liquid-Liquid SeparatorsRating: 3.5 out of 5 stars3.5/5 (3)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableFrom EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableRating: 3.5 out of 5 stars3.5/5 (22)
- Chemistry: 1001 Practice Problems For Dummies (+ Free Online Practice)From EverandChemistry: 1001 Practice Problems For Dummies (+ Free Online Practice)No ratings yet
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsFrom EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsRating: 5 out of 5 stars5/5 (3)
- Transformer: The Deep Chemistry of Life and DeathFrom EverandTransformer: The Deep Chemistry of Life and DeathRating: 4.5 out of 5 stars4.5/5 (13)
- Guidelines for Integrating Process Safety into Engineering ProjectsFrom EverandGuidelines for Integrating Process Safety into Engineering ProjectsNo ratings yet
- Monkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeFrom EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeRating: 4 out of 5 stars4/5 (9)
- Chemistry: a QuickStudy Laminated Reference GuideFrom EverandChemistry: a QuickStudy Laminated Reference GuideRating: 5 out of 5 stars5/5 (1)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeFrom EverandChemistry for Breakfast: The Amazing Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (14)
- Chemical Elements Pocket Guide: Detailed Summary of the Periodic TableFrom EverandChemical Elements Pocket Guide: Detailed Summary of the Periodic TableNo ratings yet
- Stuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldFrom EverandStuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldRating: 4 out of 5 stars4/5 (289)
- Taste: Surprising Stories and Science About Why Food Tastes GoodFrom EverandTaste: Surprising Stories and Science About Why Food Tastes GoodRating: 3 out of 5 stars3/5 (20)