You might also like
- Lymphokines: A Forum for Immunoregulatory Cell ProductsFrom EverandLymphokines: A Forum for Immunoregulatory Cell ProductsEdgar PickNo ratings yet
- Hydrolysis of InterleukinDocument12 pagesHydrolysis of InterleukinRindi GurciNo ratings yet
- Interferons and Viral Infections: AbstractDocument7 pagesInterferons and Viral Infections: Abstractjidat lawakNo ratings yet
- CPG Actua Como Adyuvante en El Cambio de La Inmunidad Con Th1 1997 CHU, ROSE S.Document9 pagesCPG Actua Como Adyuvante en El Cambio de La Inmunidad Con Th1 1997 CHU, ROSE S.Ramiro J. Rodriguez GarciaNo ratings yet
- Chest.112.6 Supplement.321SDocument9 pagesChest.112.6 Supplement.321SDuy HoangNo ratings yet
- Cytokin Nasal PoliposisDocument5 pagesCytokin Nasal PoliposisbcpNo ratings yet
- A THI-T.2 Switch Is A Critical Step in The Etiology of HIV InfectionDocument5 pagesA THI-T.2 Switch Is A Critical Step in The Etiology of HIV Infectionnik adrianiNo ratings yet
- Benevides2018 PDFDocument15 pagesBenevides2018 PDFNayche TortatoNo ratings yet
- 2011 - The Two Faces of Interferon-γ in cancerDocument12 pages2011 - The Two Faces of Interferon-γ in cancerFernando Fernández GarcíaNo ratings yet
- Il 17Document25 pagesIl 17mireya nuñezNo ratings yet
- Anti TNFDocument8 pagesAnti TNFANDONIS30No ratings yet
- 2010 - The differential activity of interferon-α subtypes is consistent among distinct target genes and cell typesDocument8 pages2010 - The differential activity of interferon-α subtypes is consistent among distinct target genes and cell typesFernando Fernández GarcíaNo ratings yet
- Colesterol: Función Biológica e Implicaciones MédicasDocument5 pagesColesterol: Función Biológica e Implicaciones MédicasJuan manuel jiménez estradaNo ratings yet
- Opal2000 PDFDocument11 pagesOpal2000 PDFKrisztián TóthNo ratings yet
- Acute TonsilitisDocument8 pagesAcute TonsilitisAdinda NadiaNo ratings yet
- Eur Grosveld 9086Document4 pagesEur Grosveld 9086carlitoseverectNo ratings yet
- Animal Models of Human DiseaseDocument9 pagesAnimal Models of Human DiseaseamamggNo ratings yet
- Yin 2009Document9 pagesYin 2009Marcos AraújoNo ratings yet
- E Lenkov 2002Document14 pagesE Lenkov 2002Matheus Tavares MartinsNo ratings yet
- ReviewDocument18 pagesReviewMUHAMMAD FARHABNo ratings yet
- RoferonDocument11 pagesRoferonisnaini fahru raziNo ratings yet
- TTNFF PDFDocument7 pagesTTNFF PDFHendroNo ratings yet
- IFN Gamma2Document8 pagesIFN Gamma2Muhammad IrawanNo ratings yet
- 2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+Document10 pages2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+顏世隆No ratings yet
- Zhu 2019Document16 pagesZhu 2019Raul ReyesNo ratings yet
- Toward Targeting in Ammasomes: Insights Into Their Regulation and ActivationDocument13 pagesToward Targeting in Ammasomes: Insights Into Their Regulation and ActivationRin ChanNo ratings yet
- Cytokine Biology - Fish Lectures: Cytokines and Their Role in The Immune SystemDocument12 pagesCytokine Biology - Fish Lectures: Cytokines and Their Role in The Immune SystemChristine QianNo ratings yet
- Interferon Gamma An Overview of Signals, Mechanisms and FunctionsDocument27 pagesInterferon Gamma An Overview of Signals, Mechanisms and FunctionsDimas ErlanggaNo ratings yet
- Type I Interferons and Their Role in Viral InfectionDocument32 pagesType I Interferons and Their Role in Viral InfectiongauhamiNo ratings yet
- Interferon GammaDocument27 pagesInterferon GammaMarcos MendezNo ratings yet
- Art 34329Document9 pagesArt 34329Maria HernándezNo ratings yet
- 58 19Document10 pages58 19Alberto RamirezNo ratings yet
- 5. Antagonist peptides of human interferon-α2b isolated from phage 2006Document7 pages5. Antagonist peptides of human interferon-α2b isolated from phage 2006Leo SirsNo ratings yet
- Cheng Et Al 2006 - Act - Il-17rDocument5 pagesCheng Et Al 2006 - Act - Il-17rNilabh RanjanNo ratings yet
- Nishikawa Et Al 2001 in The Absence of Endogenous Gamma Interferon Mice Acutely Infected With Neospora Caninum SuccumbDocument7 pagesNishikawa Et Al 2001 in The Absence of Endogenous Gamma Interferon Mice Acutely Infected With Neospora Caninum Succumbrifkyfauzan725No ratings yet
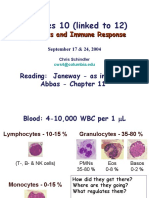
- Lectures 10 (Linked To 12) : Cytokines and Immune ResponseDocument32 pagesLectures 10 (Linked To 12) : Cytokines and Immune ResponseMac Kevin MandapNo ratings yet
- Role of Th1 and Th2 Cytokines in Immune Response To Uncomplicated Plasmodium Falciparum MalariaDocument4 pagesRole of Th1 and Th2 Cytokines in Immune Response To Uncomplicated Plasmodium Falciparum MalariaAdiebaWardaHayyaNo ratings yet
- J 1365-2249 2002 01874 XDocument8 pagesJ 1365-2249 2002 01874 XmclimacoNo ratings yet
- Alpha InterferonDocument7 pagesAlpha InterferonLAURA MARCELA BARRENECHE CALLENo ratings yet
- Alpha InterferonDocument7 pagesAlpha InterferonLAURA MARCELA BARRENECHE CALLENo ratings yet
- Role of Blimp-1 in Programing TH Effector Cells Into IL-10 ProducersDocument13 pagesRole of Blimp-1 in Programing TH Effector Cells Into IL-10 ProducerswerNo ratings yet
- Tign 1993Document7 pagesTign 1993Roberto GarceteNo ratings yet
- Punam FinalDocument13 pagesPunam FinalPratik KulkarniNo ratings yet
- Langhans 1999Document10 pagesLanghans 1999PutriPasaribuNo ratings yet
- The Effect of Antifibrotic Drug Halofugine On Th17 Cells in Concanavalin A-Induced Liver FibrosisDocument10 pagesThe Effect of Antifibrotic Drug Halofugine On Th17 Cells in Concanavalin A-Induced Liver Fibrosisa f indra pratamaNo ratings yet
- IL-10 and Its Related Superfamily Members IL-19 and IL-24 Provide Parallel Redundant Immune-Modulation in Loa Loa Infection PDFDocument9 pagesIL-10 and Its Related Superfamily Members IL-19 and IL-24 Provide Parallel Redundant Immune-Modulation in Loa Loa Infection PDFLucchéri Ndong AkomezogheNo ratings yet
- Interleukin-13 Is The Key Effector Th2 Cytokine in Ulcerative Colitis That Affects Epithelial Tight Junctions, Apoptosis, and Cell RestitutionDocument15 pagesInterleukin-13 Is The Key Effector Th2 Cytokine in Ulcerative Colitis That Affects Epithelial Tight Junctions, Apoptosis, and Cell RestitutionVinicius BalanNo ratings yet
- Aggregatibacter Actinomycetemcomitans-Induced AIM2 Inflammasome Activation Is Suppressed by Xylitol in Differentiated THP-1 MacrophagesDocument11 pagesAggregatibacter Actinomycetemcomitans-Induced AIM2 Inflammasome Activation Is Suppressed by Xylitol in Differentiated THP-1 MacrophagesRogério Ramos MarquesNo ratings yet
- Stimulated Neutrophils and Monocytes Release A Soluble FormDocument9 pagesStimulated Neutrophils and Monocytes Release A Soluble FormFco Javier Sanchez PeñaNo ratings yet
- The Role of Interleukin-1 in General PathologyDocument16 pagesThe Role of Interleukin-1 in General Pathologybaotrann.dangngocNo ratings yet
- 2010 ER Stress Induction in T CellDocument6 pages2010 ER Stress Induction in T CellRaul ReyesNo ratings yet
- Cytokine: Haiqi He, Kenneth J. Genovese, Michael H. KogutDocument7 pagesCytokine: Haiqi He, Kenneth J. Genovese, Michael H. KogutChetan Vilas JawaleNo ratings yet
- 2001gastroenterology PDFDocument12 pages2001gastroenterology PDFOUR STORYNo ratings yet
- 2001gastroenterology PDFDocument12 pages2001gastroenterology PDFOUR STORYNo ratings yet
- Burger 2019 Stressed Out About HormonesDocument10 pagesBurger 2019 Stressed Out About HormonesTriEka HeryaNo ratings yet
- Osteopontin is essential for IL‐1β production and apoptosis in peri‐implantitisDocument9 pagesOsteopontin is essential for IL‐1β production and apoptosis in peri‐implantitisBagis Emre GulNo ratings yet
- Gervaziev 2010Document8 pagesGervaziev 2010Ludmila OleninaNo ratings yet
- PAPARAN PROF. EDI DHARMANA FixDocument27 pagesPAPARAN PROF. EDI DHARMANA FixYoshe RionandoNo ratings yet
- Interleukin-17 and Its Expanding Biological Functions: ReviewDocument11 pagesInterleukin-17 and Its Expanding Biological Functions: ReviewanhiramdhaniNo ratings yet
- Il 1ra BiologyDocument29 pagesIl 1ra BiologyninaindriyaninNo ratings yet
- 0 - Application Outline For Special CategoryDocument40 pages0 - Application Outline For Special CategoryTran Trang AnhNo ratings yet
- Re Ning Procedures For The Administration of SubstancesDocument41 pagesRe Ning Procedures For The Administration of SubstancesTran Trang AnhNo ratings yet
- Unconjugated Hyperbilirubinemia in Term and Late Preterm Infants - Management - UpToDateDocument27 pagesUnconjugated Hyperbilirubinemia in Term and Late Preterm Infants - Management - UpToDateTran Trang AnhNo ratings yet
- Nelson's Pediatric Antimicrobial PDFDocument331 pagesNelson's Pediatric Antimicrobial PDFMADHANKUMAR MANICKAVELNo ratings yet
- LPS Pam3CSK 3hr 6hr 3hr 6hr Mock LPS Pam3CSK 3hr 6hr 3hr 6hr Mock Pam3CSK 3hr 6hr MockDocument5 pagesLPS Pam3CSK 3hr 6hr 3hr 6hr Mock LPS Pam3CSK 3hr 6hr 3hr 6hr Mock Pam3CSK 3hr 6hr MockTran Trang AnhNo ratings yet
- Effect of Cytokine Interplay On Macrophage Polarization During Chronic Pulmonary Infection With Cryptococcus NeoformansDocument12 pagesEffect of Cytokine Interplay On Macrophage Polarization During Chronic Pulmonary Infection With Cryptococcus NeoformansTran Trang AnhNo ratings yet
- PMN ReviewDocument10 pagesPMN ReviewPutriArifianaNo ratings yet
- Evaluation of Suspected Obstructive Sleep Apnea in Children - UpToDateDocument35 pagesEvaluation of Suspected Obstructive Sleep Apnea in Children - UpToDateTran Trang AnhNo ratings yet
- Summary Proceedings From The Neurology Group On Neonatal SeizuresDocument7 pagesSummary Proceedings From The Neurology Group On Neonatal SeizuresTran Trang AnhNo ratings yet
- E0004 FullDocument6 pagesE0004 FullBaharudin WahyuNo ratings yet
- CPAP For Pediatric Obstructive Sleep Apnea - UpToDateDocument18 pagesCPAP For Pediatric Obstructive Sleep Apnea - UpToDateTran Trang AnhNo ratings yet
- Cardiovascular Consequences of Obstructive Sleep Apnea in Children - UpToDateDocument16 pagesCardiovascular Consequences of Obstructive Sleep Apnea in Children - UpToDateTran Trang AnhNo ratings yet
- Management of Obstructive Sleep Apnea in Children - UpToDateDocument21 pagesManagement of Obstructive Sleep Apnea in Children - UpToDateTran Trang AnhNo ratings yet
- Mechanisms and Predisposing Factors For Sleep-Related Breathing Disorders in Children - UpToDateDocument30 pagesMechanisms and Predisposing Factors For Sleep-Related Breathing Disorders in Children - UpToDateTran Trang AnhNo ratings yet
- (Special Edition) 20 Things To Do in The Castle Town of KanazawaDocument12 pages(Special Edition) 20 Things To Do in The Castle Town of KanazawaTran Trang AnhNo ratings yet
- Consensus Conference Methodology: British Journal of Obstetrics and GynaecologyDocument2 pagesConsensus Conference Methodology: British Journal of Obstetrics and GynaecologyTran Trang AnhNo ratings yet
- BMDM - Bone Marrow-Derived Macrophage ProductionDocument6 pagesBMDM - Bone Marrow-Derived Macrophage ProductionTran Trang AnhNo ratings yet
- Grading Quality of Evidence and Strength ofDocument5 pagesGrading Quality of Evidence and Strength ofArkipus PamuttuNo ratings yet
- 1ielts Support Tools 2018Document28 pages1ielts Support Tools 2018Bacem FarhatNo ratings yet
- Grading Guide: Why Uptodate? Product Editorial Subscription Options Subscribe Uptodate®Document17 pagesGrading Guide: Why Uptodate? Product Editorial Subscription Options Subscribe Uptodate®Tran Trang AnhNo ratings yet
- Terminologie: Méningite & EncéphaliteDocument10 pagesTerminologie: Méningite & EncéphaliteTran Trang AnhNo ratings yet
- Fever: in Children: ShouldDocument4 pagesFever: in Children: ShouldTran Trang AnhNo ratings yet
- 690Document1 page690Tran Trang AnhNo ratings yet
- Masondarmawan 2016 English-Version PDFDocument32 pagesMasondarmawan 2016 English-Version PDFTran Trang AnhNo ratings yet
- Integration Stability of Shbsag-Multi Expression Cassettes in Pichia Pastoris Gs115 During Methanol InductionDocument7 pagesIntegration Stability of Shbsag-Multi Expression Cassettes in Pichia Pastoris Gs115 During Methanol InductionAlex MaxNo ratings yet
- Passmrcog Micro2016Document130 pagesPassmrcog Micro2016Cwali MohamedNo ratings yet
- 13 KuzmanDocument7 pages13 KuzmanSinhad Dadi GolubovicNo ratings yet
- Meril COVID-19 One-Step RT-PCR KitDocument2 pagesMeril COVID-19 One-Step RT-PCR KitAnkit RanaNo ratings yet
- Physical and Chemical Methods of SterilizationDocument27 pagesPhysical and Chemical Methods of SterilizationDanny Mel ManglaylayNo ratings yet
- Microbiology and Parasitology Presentation by W KakumuraDocument14 pagesMicrobiology and Parasitology Presentation by W KakumurahamiltonNo ratings yet
- Feline PanleukopeniaDocument10 pagesFeline PanleukopeniaDhea SalsabilaNo ratings yet
- TA Brown Chapter 2 NotesDocument7 pagesTA Brown Chapter 2 Notesom prakashNo ratings yet
- Bouchentouf and MissoumDocument13 pagesBouchentouf and MissoumSilvia MariluzNo ratings yet
- Summer Camp Guidance 2021: Health and Safety GuidanceDocument6 pagesSummer Camp Guidance 2021: Health and Safety GuidanceViens DiviNo ratings yet
- VERI-Q™ PCR 316 Detection Platform: COVID-19Document2 pagesVERI-Q™ PCR 316 Detection Platform: COVID-19Smithesh TpNo ratings yet
- Analisis Dinamik Model SIR Pada Kasus Penyebaran Penyakit Corona Virus Disease-19 (COVID-19)Document8 pagesAnalisis Dinamik Model SIR Pada Kasus Penyebaran Penyakit Corona Virus Disease-19 (COVID-19)DHEA AMELIANo ratings yet
- Informal Letter DengueDocument2 pagesInformal Letter DengueIrsyad IzzatNo ratings yet
- Potong PDFDocument24 pagesPotong PDFMaryo Osan SinaNo ratings yet
- Current Event Analysis Step 2 GAVIN KDocument1 pageCurrent Event Analysis Step 2 GAVIN KGAVIN KURNIAWANNo ratings yet
- Second Thoughts About Viruses Vaccines and The HIV Part 1 Robert.O.YoungDocument8 pagesSecond Thoughts About Viruses Vaccines and The HIV Part 1 Robert.O.YoungDavidNo ratings yet
- D-37/1, TTC MIDC, Turbhe, Navi Mumbai-400 703: Processed At: ThyrocareDocument3 pagesD-37/1, TTC MIDC, Turbhe, Navi Mumbai-400 703: Processed At: ThyrocareKAMAL KANTNo ratings yet
- MicrobiologyPharm D Notes For Spiral BindingDocument282 pagesMicrobiologyPharm D Notes For Spiral BindingLini Sara KoshyNo ratings yet
- COVID-19 Abortion Policy LetterDocument5 pagesCOVID-19 Abortion Policy LetterMinnesota House Republicans100% (2)
- Potential Conflicts of Interest. All Authors:: Clinical Infectious Diseases 2018 66 (8) :1316-7Document2 pagesPotential Conflicts of Interest. All Authors:: Clinical Infectious Diseases 2018 66 (8) :1316-7Chikondi TemboNo ratings yet
- Short Story COVID-19Document3 pagesShort Story COVID-19Kim SaysonNo ratings yet
- University of Leeds Dissertation SubmissionDocument5 pagesUniversity of Leeds Dissertation SubmissionPaperHelpCanada100% (1)
- Article-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and SurvivalDocument25 pagesArticle-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and Survivalcarlos ArozamenaNo ratings yet
- Pathology MCQ - Liver and Biliary Tract PDFDocument5 pagesPathology MCQ - Liver and Biliary Tract PDFCHRISTOPHER OWUSU ASARENo ratings yet
- 03apr - HT Mumbai PDFDocument14 pages03apr - HT Mumbai PDFpannapurohitNo ratings yet
- A Antimicrobial Susceptibility TestingDocument13 pagesA Antimicrobial Susceptibility TestingkurfaNo ratings yet
- Group 10 - Stem 11 ST - DominicDocument34 pagesGroup 10 - Stem 11 ST - Dominicchristine ancheta100% (1)
- Acute Encephalitis SyndromeDocument116 pagesAcute Encephalitis SyndromePrateek Kumar PandaNo ratings yet
- Tools and Resources - Cited-AntibodiesDocument24 pagesTools and Resources - Cited-AntibodiesAnkit GuptaNo ratings yet
- 2018 MTE GIVD Classification 2018 Rev01Document14 pages2018 MTE GIVD Classification 2018 Rev01Dr.Sivakumar SelvarajNo ratings yet