You might also like
- The Stock-Feeder's Manual the chemistry of food in relation to the breeding and feeding of live stockFrom EverandThe Stock-Feeder's Manual the chemistry of food in relation to the breeding and feeding of live stockNo ratings yet
- Pared BacteriasDocument18 pagesPared BacteriasenadesNo ratings yet
- Gross, Biocheem.: A On To Sources To Our Diffusion Kinetics.3 Ture Stances AsDocument8 pagesGross, Biocheem.: A On To Sources To Our Diffusion Kinetics.3 Ture Stances Asellla888No ratings yet
- Mixed SedimentDocument15 pagesMixed SedimentprashmceNo ratings yet
- Asistensi BakterDocument15 pagesAsistensi BakterAwaliatun Nur AzizahNo ratings yet
- Sporulation of Aspergillus Niger and AspergillusDocument10 pagesSporulation of Aspergillus Niger and AspergillusGanesh KashinathNo ratings yet
- Isolation A N D Characteristics Hafiz Plate References S. DDocument9 pagesIsolation A N D Characteristics Hafiz Plate References S. DKen WongNo ratings yet
- 166949a0 PDFDocument1 page166949a0 PDFKahkashan AhmedNo ratings yet
- Cholesterol-Reducing Bacterium From Human FecesDocument8 pagesCholesterol-Reducing Bacterium From Human FecesfrankcarnationNo ratings yet
- The Occurrence of Chitinase in Some Bacteria: Gen. Microbiol. 14Document9 pagesThe Occurrence of Chitinase in Some Bacteria: Gen. Microbiol. 14Illani Zuraihah IbrahimNo ratings yet
- T H e Nutrient Content of High and Low Quality Fresh Eggs. I. Total Solids, Total Nitrogen, 1 (-) and TryptophaneDocument6 pagesT H e Nutrient Content of High and Low Quality Fresh Eggs. I. Total Solids, Total Nitrogen, 1 (-) and TryptophaneYoselin GomezNo ratings yet
- JB 82 4 582-588 1961Document7 pagesJB 82 4 582-588 1961laasyagudiNo ratings yet
- TermoresistenciaDocument10 pagesTermoresistenciaIvan SanchezNo ratings yet
- Acid Other MusDocument9 pagesAcid Other MusRoberto CastellanosNo ratings yet
- Identification of Strains Using The API System: BacillusDocument12 pagesIdentification of Strains Using The API System: BacillusMohamed Wassim M'henniNo ratings yet
- 1978 Chesson, The Maceration of Linen Flax Under Anaerobic ConditionsDocument12 pages1978 Chesson, The Maceration of Linen Flax Under Anaerobic Conditionsm222000No ratings yet
- 1 s2.0 0740002087900037 MainDocument11 pages1 s2.0 0740002087900037 MainLarissa RodriguesNo ratings yet
- Growth Kinetics of Thiobacillus Denitrificans in A PDFDocument9 pagesGrowth Kinetics of Thiobacillus Denitrificans in A PDFPatrick Antony Bent BowieNo ratings yet
- Antimicrobial Properties of Salt Natural CasingsDocument6 pagesAntimicrobial Properties of Salt Natural CasingsdesmadradorNo ratings yet
- Isolation, Identification and Characterization of Salmonella From DuckDocument6 pagesIsolation, Identification and Characterization of Salmonella From DuckAmira Fathini AzmanNo ratings yet
- Fermentação de Glicose Por ChlorellaDocument8 pagesFermentação de Glicose Por ChlorellapaulavonNo ratings yet
- Designacion de H37Rv Como Neotipo de MTBDocument8 pagesDesignacion de H37Rv Como Neotipo de MTBSamantha YongNo ratings yet
- Pnas00161 0050Document8 pagesPnas00161 0050Jonathan MoralesNo ratings yet
- Desarrollo y Dinamica Poblacional TisbeDocument15 pagesDesarrollo y Dinamica Poblacional TisbepablofedericciNo ratings yet
- Enterococcus and Lactobacillus Contamination of Raw Milk in A Farm Dairy EnvironmentDocument9 pagesEnterococcus and Lactobacillus Contamination of Raw Milk in A Farm Dairy Environmentbibliomed08No ratings yet
- 12 Kligler Iron AgarDocument3 pages12 Kligler Iron AgarOsama BakheetNo ratings yet
- Physiological Studies of Subtropical Mangrove ThraustochytridsDocument8 pagesPhysiological Studies of Subtropical Mangrove ThraustochytridsIsnaini PrihatiningsihNo ratings yet
- 2685 FullDocument8 pages2685 FullAndréRochaNo ratings yet
- The Influence of Medium Composition On The Growth and Swarming of ProteusDocument11 pagesThe Influence of Medium Composition On The Growth and Swarming of ProteusfarikoNo ratings yet
- AN165MDocument7 pagesAN165MNOYNo ratings yet
- Morphogenesis Arthrobacter Crystallopoietes: C. ANDDocument9 pagesMorphogenesis Arthrobacter Crystallopoietes: C. ANDRyan RachmawanNo ratings yet
- Plant Growth Substances Produced by Micro-Organisms Soil and RhizosphereDocument9 pagesPlant Growth Substances Produced by Micro-Organisms Soil and RhizosphereLAURA DANIELA VALDERRAMA TORRESNo ratings yet
- Carbon Va NitoDocument11 pagesCarbon Va NitoHoang AnhNo ratings yet
- Food Microbiology: Amit Pal, Douglas L. MarshallDocument3 pagesFood Microbiology: Amit Pal, Douglas L. MarshallHooi Jai BoonNo ratings yet
- The Use of Elements As A Substitute For Biomass in Toxicokinetic Studies in Small OrganismsDocument7 pagesThe Use of Elements As A Substitute For Biomass in Toxicokinetic Studies in Small OrganismsBruno GomesNo ratings yet
- Indctn Reprssn CELLULASEDocument13 pagesIndctn Reprssn CELLULASEfragarfloNo ratings yet
- Microbiological Survey of Georgia Poultry LitterDocument9 pagesMicrobiological Survey of Georgia Poultry Litteredit16No ratings yet
- B. GinsenosoiliDocument5 pagesB. GinsenosoiliEl-Agamy ProbeNo ratings yet
- Studies On TO Toxins of Clostridium Botulinum A Simplified ProcedureDocument6 pagesStudies On TO Toxins of Clostridium Botulinum A Simplified ProcedureAustinNo ratings yet
- 2014 Skaloud Et Al PlanktochlorellaDocument10 pages2014 Skaloud Et Al Planktochlorellanita prmtasariNo ratings yet
- Sublethal Stress in Escherichia Coli A Function of SalinitytDocument6 pagesSublethal Stress in Escherichia Coli A Function of SalinitytgiuseppegnrNo ratings yet
- Isolation and Identification of Iron-Reducing Bacteria From Gley SoilsDocument13 pagesIsolation and Identification of Iron-Reducing Bacteria From Gley SoilsMel EsparzaNo ratings yet
- A-heavy-metal-tolerant-novel-bacterium,-Bacillus-malikii-sp.-nov.,-isolated-from-tannery-effluent-wastewater_2015_Antonie-van-Leeuwenhoek,-International-Journal-of-General-and-Molecular-Microbiology.pdfDocument12 pagesA-heavy-metal-tolerant-novel-bacterium,-Bacillus-malikii-sp.-nov.,-isolated-from-tannery-effluent-wastewater_2015_Antonie-van-Leeuwenhoek,-International-Journal-of-General-and-Molecular-Microbiology.pdfCristian SalgadoNo ratings yet
- Microbial Contamination of Pharmaceutical PreparationsDocument6 pagesMicrobial Contamination of Pharmaceutical PreparationsReiamury AzraqNo ratings yet
- Poultry FeedDocument7 pagesPoultry FeedRahul AmbawataNo ratings yet
- Qualidade PescadoDocument15 pagesQualidade PescadoAndré CampeloNo ratings yet
- Nakas, J. P., Et Al. "System Development For Linked-Fermentation Production of Solvents From Algal Biomass."Document7 pagesNakas, J. P., Et Al. "System Development For Linked-Fermentation Production of Solvents From Algal Biomass."Mafalda MarquesNo ratings yet
- Scientific Proceedings: Detroit, May 21 24, 1946Document22 pagesScientific Proceedings: Detroit, May 21 24, 1946Veneta GizdakovaNo ratings yet
- Effect of Sulfonamides and Thioureas On The Thyroid Gland and Basal MetabolismDocument25 pagesEffect of Sulfonamides and Thioureas On The Thyroid Gland and Basal MetabolismM SaqibNo ratings yet
- Appl. Microbiol.-1971-Kim-581-7Document7 pagesAppl. Microbiol.-1971-Kim-581-7gmanju207No ratings yet
- 1960 - Amino Acid Decarboxylases in A PseudomonaDocument7 pages1960 - Amino Acid Decarboxylases in A PseudomonaGiancarlo GHNo ratings yet
- The Growth of Penicillium Roqueforti On Synthetic MediaDocument14 pagesThe Growth of Penicillium Roqueforti On Synthetic MediaFranco VeglianiNo ratings yet
- Mushroom Culture Media PDFDocument11 pagesMushroom Culture Media PDFAnonymous z5pzOM100% (1)
- Isolation and Identification of Avibacterium Paragallinarum, The Causal Agent of Infectious Coryza (IC) From Layer Chickens in BangladeshDocument10 pagesIsolation and Identification of Avibacterium Paragallinarum, The Causal Agent of Infectious Coryza (IC) From Layer Chickens in BangladeshTaufiqAhmadFariziNo ratings yet
- Fungal Spore Germination PercentageDocument1 pageFungal Spore Germination PercentageArjun ShamNo ratings yet
- Isolation and Characterization of Acetobacter Aceti From Rotten PapayaDocument8 pagesIsolation and Characterization of Acetobacter Aceti From Rotten PapayasivabioteckNo ratings yet
- Isolation Actinomycetes ANDRIANODocument7 pagesIsolation Actinomycetes ANDRIANOBerenice AndrianoNo ratings yet
- Ijs 0 63682-0Document6 pagesIjs 0 63682-0ehab homoudNo ratings yet
- Biochem - Pityrosporum OvaleDocument7 pagesBiochem - Pityrosporum OvaleJuliana SoaresNo ratings yet
- Degradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusDocument7 pagesDegradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusMuhammad UmerNo ratings yet
- Ac Bihar - 55Document1 pageAc Bihar - 55Kamran ZahirNo ratings yet
- Booth Under Narpatganj Block, Araria District: Prepared byDocument1 pageBooth Under Narpatganj Block, Araria District: Prepared byKamran ZahirNo ratings yet
- Ucg 1106 Gsi PDFDocument68 pagesUcg 1106 Gsi PDFbandita samalNo ratings yet
- Booth Under Sikti Block, Araria District: IndexDocument1 pageBooth Under Sikti Block, Araria District: IndexKamran ZahirNo ratings yet
- Araria District: A Geographical Out Look: Chapter-2Document30 pagesAraria District: A Geographical Out Look: Chapter-2Kamran ZahirNo ratings yet
- Neet Sample Paper Biology Question Paper: IndiaDocument13 pagesNeet Sample Paper Biology Question Paper: IndiaKamran ZahirNo ratings yet
- Outcome of Scoping Process Since March Draft Guiding Principles Building BlocksDocument22 pagesOutcome of Scoping Process Since March Draft Guiding Principles Building BlocksKamran ZahirNo ratings yet
- 59 4 181 PDFDocument13 pages59 4 181 PDFKamran ZahirNo ratings yet
- Academic Planning 2013-2014: Syllabus - Class IDocument24 pagesAcademic Planning 2013-2014: Syllabus - Class IKamran ZahirNo ratings yet
- Academic Planning 2013-2014: Syllabus - Class IDocument24 pagesAcademic Planning 2013-2014: Syllabus - Class IKamran ZahirNo ratings yet
- 051 - SIKTI: Morang (Nepal) Sunsari (Nepal)Document1 page051 - SIKTI: Morang (Nepal) Sunsari (Nepal)Kamran ZahirNo ratings yet
- Pubg Game PDFDocument1 pagePubg Game PDFKamran ZahirNo ratings yet
- 051 - SIKTI: Morang (Nepal) Sunsari (Nepal)Document1 page051 - SIKTI: Morang (Nepal) Sunsari (Nepal)Kamran ZahirNo ratings yet
- Od118129413217650000 PDFDocument1 pageOd118129413217650000 PDFKamran ZahirNo ratings yet
- Staphylococci and Their Classification: by Constance Shaw, Jennifer M. StittDocument14 pagesStaphylococci and Their Classification: by Constance Shaw, Jennifer M. StittKamran ZahirNo ratings yet
- IJMHAPUBGImpacts India Cases 2019Document6 pagesIJMHAPUBGImpacts India Cases 2019Debashis BhaskarNo ratings yet
- Od118129413217650000 PDFDocument1 pageOd118129413217650000 PDFKamran ZahirNo ratings yet
- Model Questions Mathematics: FX XXRDocument37 pagesModel Questions Mathematics: FX XXRdivakar sumanNo ratings yet
- Sup 0A 0033Document12 pagesSup 0A 0033Kamran ZahirNo ratings yet
- Atomic StructureDocument37 pagesAtomic Structurekishangopi123100% (1)
- 000 IntroAndOverviewDocument21 pages000 IntroAndOverviewThế Phương LưuNo ratings yet
- Od118129413217650000 PDFDocument1 pageOd118129413217650000 PDFKamran ZahirNo ratings yet
- Clinical Microbiology QuestionsDocument18 pagesClinical Microbiology QuestionsArianne Joy C. TamarayNo ratings yet
- Systemic Mycoses: Cold Mold Heat YeastDocument4 pagesSystemic Mycoses: Cold Mold Heat YeastHanunNo ratings yet
- Identification of Potentially Pathogenic Bacteria From Tilapia (Oreochromis Niloticus) and Channel Catfish (Clarias Batrachus) Culture in Samarinda, East Kalimantan, IndonesiaDocument9 pagesIdentification of Potentially Pathogenic Bacteria From Tilapia (Oreochromis Niloticus) and Channel Catfish (Clarias Batrachus) Culture in Samarinda, East Kalimantan, IndonesianurullahfatihNo ratings yet
- Putting On and Removing Ppe Diagram - March 2020Document2 pagesPutting On and Removing Ppe Diagram - March 2020Perdana DialisisNo ratings yet
- Principle General Information PreparationDocument2 pagesPrinciple General Information PreparationHaidar HanifNo ratings yet
- Rd-15 Bartholin Cyst and Abscess 10-12Document2 pagesRd-15 Bartholin Cyst and Abscess 10-12Chong Hui ChekNo ratings yet
- Bacterial Vaginosis Treatment - UpToDateDocument12 pagesBacterial Vaginosis Treatment - UpToDateAlex Esteban Espinoza CevallosNo ratings yet
- Cleaning, Disinfecting, Sanitizing Difference FlyerDocument1 pageCleaning, Disinfecting, Sanitizing Difference FlyerFizz FirdausNo ratings yet
- FILARIASISDocument31 pagesFILARIASISArun Shree RNo ratings yet
- Graduating Students Consent FormDocument1 pageGraduating Students Consent FormZack ClaymanNo ratings yet
- Study Notes For Sexually Transmitted DiseasesDocument4 pagesStudy Notes For Sexually Transmitted DiseasesPrince K. TaileyNo ratings yet
- Non-Enteric Gastrointestinal PathogensDocument11 pagesNon-Enteric Gastrointestinal PathogensOrhan AsdfghjklNo ratings yet
- Demam Tifoid FTKP Bpjs Hotel ClarionDocument68 pagesDemam Tifoid FTKP Bpjs Hotel ClarionPeriyent Liveryent VozzervodNo ratings yet
- Susceptible, Infected, Recovered: The SIR Model of An EpidemicDocument16 pagesSusceptible, Infected, Recovered: The SIR Model of An Epidemicmirast91No ratings yet
- Assessment of Enzymatic Cleaning Agents and Disinfectants Against Bacterial BiofilmsDocument10 pagesAssessment of Enzymatic Cleaning Agents and Disinfectants Against Bacterial BiofilmsSoumya Saswat PandaNo ratings yet
- Test Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner BairDocument35 pagesTest Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner Bairshriekacericg31u3100% (37)
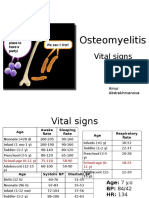
- PBL 2 - OsteomyelitisDocument10 pagesPBL 2 - OsteomyelitisAinur AbdrakhmanovaNo ratings yet
- CH4 NotesDocument10 pagesCH4 NotesShane TamilNo ratings yet
- Gram Negative RodsDocument23 pagesGram Negative RodsYasir KareemNo ratings yet
- Lacto BacillusDocument10 pagesLacto BacillusLucasNo ratings yet
- Hospital Management: Bacterial InfectionDocument21 pagesHospital Management: Bacterial Infectionapadanainst7372No ratings yet
- Molecular Diagnostics: Report Status - Final Test Name Result Biological Ref. Interval UnitDocument3 pagesMolecular Diagnostics: Report Status - Final Test Name Result Biological Ref. Interval UnitARVIND SINGHNo ratings yet
- HSB Pneumonia Antibiotic AlgorithmDocument4 pagesHSB Pneumonia Antibiotic AlgorithmDr.Senthil KumarNo ratings yet
- PHBIOSCI 5B - ACTIVITY #1 - Basic Laboratory Techniques For Isolation, Cultivation, and Cultural Characterization of Microorganisms (SUMMER 2022)Document1 pagePHBIOSCI 5B - ACTIVITY #1 - Basic Laboratory Techniques For Isolation, Cultivation, and Cultural Characterization of Microorganisms (SUMMER 2022)Shopifyy ClothingNo ratings yet
- Endoftalmitis EndogenaDocument1 pageEndoftalmitis EndogenaAlexander Fabián Rodríguez DíazNo ratings yet
- 1.1 Identify The Differences Between Bacteria, Viruses, Fungi and ParasitesDocument4 pages1.1 Identify The Differences Between Bacteria, Viruses, Fungi and ParasitesgemmaNo ratings yet
- MLTDocument34 pagesMLTJai MurugeshNo ratings yet
- Bubonic PlagueDocument15 pagesBubonic Plagueapi-283967550No ratings yet
- Rosa Urdaneta 150 Sunny Isles BLVD Miami, FL 33160: Test ReportDocument2 pagesRosa Urdaneta 150 Sunny Isles BLVD Miami, FL 33160: Test ReportRosaNo ratings yet
- Microbes NotesDocument22 pagesMicrobes Notesapi-218511741100% (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4 out of 5 stars4/5 (5)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)