You might also like
- PART 2 Coran Pediatric Surgery, 7th Ed PDFDocument350 pagesPART 2 Coran Pediatric Surgery, 7th Ed PDFRamona Cerasela Petreaca100% (4)
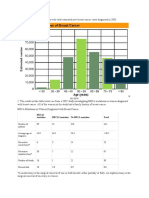
- Brca1 Mutation BRCA2 Mutation No BRCA Mutation Total: © 2011 FLVSDocument3 pagesBrca1 Mutation BRCA2 Mutation No BRCA Mutation Total: © 2011 FLVSJasmine VeraNo ratings yet
- Non Epithelial Ovarian CancerDocument18 pagesNon Epithelial Ovarian CancerDR RISKA WAHYUNo ratings yet
- No Epitelial Ovarion Cancer 2018Document18 pagesNo Epitelial Ovarion Cancer 2018palomazul007No ratings yet
- Cancers: Therapeutic Challenges For Cisplatin-Resistant Ovarian Germ Cell TumorsDocument14 pagesCancers: Therapeutic Challenges For Cisplatin-Resistant Ovarian Germ Cell Tumorsatikha apriliaNo ratings yet
- Article G CTDocument9 pagesArticle G CTLaura Paola UrquizoNo ratings yet
- Current Controversies in The Management of Germ Cell Ovarian TumoursDocument7 pagesCurrent Controversies in The Management of Germ Cell Ovarian Tumoursatikha apriliaNo ratings yet
- Vazquez2013 PDFDocument7 pagesVazquez2013 PDFatikha apriliaNo ratings yet
- Pathogenesis of Testicular Germ Cell Tumours: Leendert H. J. Looijenga and J. Wolter OosterhuisDocument11 pagesPathogenesis of Testicular Germ Cell Tumours: Leendert H. J. Looijenga and J. Wolter OosterhuisLatansa DinaNo ratings yet
- Human Germ Cell Tumors Are Developmental Cancers: Impact of Epigenetics On Pathobiology and ClinicDocument28 pagesHuman Germ Cell Tumors Are Developmental Cancers: Impact of Epigenetics On Pathobiology and Clinicarcobaleno589No ratings yet
- 624 PDFDocument9 pages624 PDFTri AryantiNo ratings yet
- Jurnal Kanker TestisDocument9 pagesJurnal Kanker TestisDiatni FibriNo ratings yet
- Testis CancerDocument9 pagesTestis CancerMuhammad Tawfiq ZamriNo ratings yet
- Glioblastoma Stem-Like Cells Approaches For Isolation and CharacterizationDocument19 pagesGlioblastoma Stem-Like Cells Approaches For Isolation and CharacterizationMariano PerezNo ratings yet
- Sacrococcygeal Teratoma A Tumor at The Center of EmbryogenesisDocument8 pagesSacrococcygeal Teratoma A Tumor at The Center of EmbryogenesisLuis Ruelas SanchezNo ratings yet
- Jurnal Tumor Metastasis Paru Hepar Dari PancreasDocument15 pagesJurnal Tumor Metastasis Paru Hepar Dari PancreaslucyishakNo ratings yet
- Diagnostics 12 02050 v2Document23 pagesDiagnostics 12 02050 v2Alin VasileNo ratings yet
- 331-Book Chapter-3614-2-10-20210406Document20 pages331-Book Chapter-3614-2-10-20210406Yolla GitamayaNo ratings yet
- CX3CR1 Acts As A Protective Biomarker in The Tumor Microenvironment of Colorectal CancerDocument16 pagesCX3CR1 Acts As A Protective Biomarker in The Tumor Microenvironment of Colorectal CancerIsmaNo ratings yet
- Cancers 12 00611Document17 pagesCancers 12 00611IRINA BRANNo ratings yet
- Cell Bio CriticDocument24 pagesCell Bio CriticRetchel Cadayong NavalesNo ratings yet
- FDG PET CT in Testicular CancerDocument4 pagesFDG PET CT in Testicular CancerlkitkatgirlNo ratings yet
- Endometrialcancer: Obesity, Genetics, and Targeted AgentsDocument17 pagesEndometrialcancer: Obesity, Genetics, and Targeted AgentsEftychia GkikaNo ratings yet
- Article 5 POCDocument16 pagesArticle 5 POCmariimaria670No ratings yet
- Anatomy and Pathology of Testicular Tumors - UpToDateDocument26 pagesAnatomy and Pathology of Testicular Tumors - UpToDateBhargav YagnikNo ratings yet
- 10.1016/j.humpath.2016.08.002: Human PathologyDocument50 pages10.1016/j.humpath.2016.08.002: Human PathologyAdryansyah ChaniagoNo ratings yet
- Medsci 06 00031 PDFDocument113 pagesMedsci 06 00031 PDFCarolina VillalobosNo ratings yet
- Clsificacion 3Document10 pagesClsificacion 3Paulo Cesar Castañeda RuizNo ratings yet
- Critical Reviews in Oncology / HematologyDocument9 pagesCritical Reviews in Oncology / HematologyNadya MagfiraNo ratings yet
- Kjco 14 2 83Document6 pagesKjco 14 2 83Antonio GligorievskiNo ratings yet
- Genetics GCTDocument10 pagesGenetics GCTEvelynππ θσυNo ratings yet
- Extragonadal Germ Cell Tumors: H.-J. SchmollDocument8 pagesExtragonadal Germ Cell Tumors: H.-J. SchmollAshutoshNo ratings yet
- Thyroid Cancer Metabolism A Review 2167 7948 1000200Document7 pagesThyroid Cancer Metabolism A Review 2167 7948 1000200yukii celloNo ratings yet
- Selle2015 PDFDocument12 pagesSelle2015 PDFatikha apriliaNo ratings yet
- Pediatric Thyroid Cancer 2017Document20 pagesPediatric Thyroid Cancer 2017Alin VázquezNo ratings yet
- Rodriguez-Galindo2005, Ribeiro, BonaldDocument9 pagesRodriguez-Galindo2005, Ribeiro, BonaldLeonardo Müller RodriguesNo ratings yet
- Classi Fication of Endometrial CarcinomaDocument23 pagesClassi Fication of Endometrial CarcinomaKelly F RuizNo ratings yet
- Roylance 1997Document8 pagesRoylance 1997barti koksNo ratings yet
- 10.1007@s12094 010 0599 0Document5 pages10.1007@s12094 010 0599 0ZULMA MADELINE LIBNY GUTARRA TICANo ratings yet
- Bindea Et Al. - 2011 - The Prognostic Impact of Anti-Cancer Immune Response A Novel Classification of Cancer Patients PDFDocument6 pagesBindea Et Al. - 2011 - The Prognostic Impact of Anti-Cancer Immune Response A Novel Classification of Cancer Patients PDFflashjetNo ratings yet
- Germ Cell Tumors Ovarium: Poppy M. LintongDocument12 pagesGerm Cell Tumors Ovarium: Poppy M. Lintongvytania oktariNo ratings yet
- Clinicopathological Study of Metallothionein Immunohistochemical Expression, in Benign, Borderline and Malignant Ovarian Epithelial TumorsDocument7 pagesClinicopathological Study of Metallothionein Immunohistochemical Expression, in Benign, Borderline and Malignant Ovarian Epithelial TumorsNajmul HasanNo ratings yet
- Cancers Make Their Own Luck - Theories of Cancer OriginsDocument15 pagesCancers Make Their Own Luck - Theories of Cancer Originszhe zhNo ratings yet
- Molecular Carcinogenesis Endometrium PDFDocument7 pagesMolecular Carcinogenesis Endometrium PDFMelati HasnailNo ratings yet
- The Importance of Genomic Copy Number Changes in The Prognosis of Glioblastoma MultiformeDocument7 pagesThe Importance of Genomic Copy Number Changes in The Prognosis of Glioblastoma MultiformeALINo ratings yet
- Tumori Testisa PDFDocument14 pagesTumori Testisa PDFVuk MilutinovićNo ratings yet
- Association of Glutathione S-TDocument6 pagesAssociation of Glutathione S-TAinunNo ratings yet
- Potential Risk Factors of Glioblastoma Multiforme in Greek Adults A Case Control StudyDocument17 pagesPotential Risk Factors of Glioblastoma Multiforme in Greek Adults A Case Control StudyAthenaeum Scientific PublishersNo ratings yet
- JCR 23 007Document13 pagesJCR 23 007yakalapunagarajuNo ratings yet
- Imun PDFDocument4 pagesImun PDFLeyni SabatiniNo ratings yet
- G Path-NeoplasiaDocument60 pagesG Path-Neoplasiachouchou124No ratings yet
- Recurrent Driver Mutations in Benign TumorsDocument10 pagesRecurrent Driver Mutations in Benign Tumorsdanielnobre47570No ratings yet
- Research Essay Final - Aidan HargettDocument10 pagesResearch Essay Final - Aidan Hargettapi-559292279No ratings yet
- Cancer Is A Functional Repair TissueDocument6 pagesCancer Is A Functional Repair TissueFilipos ConstantinNo ratings yet
- Cpho023 01 03Document11 pagesCpho023 01 03Kyung-Nam KohNo ratings yet
- Pineal Region Tumors PDFDocument15 pagesPineal Region Tumors PDFhimadriNo ratings yet
- BCR 554Document6 pagesBCR 554Yuanita MardastutiNo ratings yet
- The Mechanisms of Cancer-Associated Thrombosis: Anna Falanga, Marina Marchetti, Laura RussoDocument4 pagesThe Mechanisms of Cancer-Associated Thrombosis: Anna Falanga, Marina Marchetti, Laura RussoThomaz DamianiNo ratings yet
- Arar0332 Jco 2021 DR RaulDocument14 pagesArar0332 Jco 2021 DR RaulLeonardo Müller RodriguesNo ratings yet
- FigoDocument11 pagesFigoPraja Putra AdnyanaNo ratings yet
- Human Germ Cell Tumors Are Developmental Cancers: Impact of Epigenetics On Pathobiology and ClinicDocument28 pagesHuman Germ Cell Tumors Are Developmental Cancers: Impact of Epigenetics On Pathobiology and Clinicarcobaleno589No ratings yet
- Jkns 57 315Document8 pagesJkns 57 315arcobaleno589No ratings yet
- HHS Public Access: Primary Central Nervous System Germ Cell Tumors: A Review and UpdateDocument15 pagesHHS Public Access: Primary Central Nervous System Germ Cell Tumors: A Review and Updatearcobaleno589No ratings yet
- Case Report: Intracranial Multiple Germ Cell Tumors: A Case Report and Review of LiteratureDocument6 pagesCase Report: Intracranial Multiple Germ Cell Tumors: A Case Report and Review of Literaturearcobaleno589No ratings yet
- Hypertension in Adults Diagnosis and Management PDF 66141722710213 PDFDocument40 pagesHypertension in Adults Diagnosis and Management PDF 66141722710213 PDFarcobaleno589No ratings yet
- Hemostasis and Blood CoagulationDocument12 pagesHemostasis and Blood Coagulationarcobaleno589No ratings yet
- ARDSDocument6 pagesARDSarcobaleno589No ratings yet
- 2,4-Diaminoanisole and Its Salts: 1.1 Chemical and Physical DataDocument25 pages2,4-Diaminoanisole and Its Salts: 1.1 Chemical and Physical DatakhoachiNo ratings yet
- Distribution and Habitat: Jasminum Sambac Jasminum Mesnyi Jasminum AngustifoliumDocument1 pageDistribution and Habitat: Jasminum Sambac Jasminum Mesnyi Jasminum AngustifoliumpriyankaNo ratings yet
- Bryophytes A Potential Source of AntioxidantsDocument13 pagesBryophytes A Potential Source of AntioxidantsROBERTO MIRANDA CALDASNo ratings yet
- Lif101a 111a FCHDocument1 pageLif101a 111a FCHnemoj23473No ratings yet
- Unit 13 Cleavage and GastrulationDocument26 pagesUnit 13 Cleavage and GastrulationFERHAEEZA KALAYAKANNo ratings yet
- 13-12th Group B Half Book Bio Chap 15+25+27,19+21Document4 pages13-12th Group B Half Book Bio Chap 15+25+27,19+21Raees Ali KhanNo ratings yet
- Clinical Review, December 16, 2017 - LUXTURNADocument94 pagesClinical Review, December 16, 2017 - LUXTURNAFatema RavatNo ratings yet
- 1 s2.0 S0753332220311896 MainDocument12 pages1 s2.0 S0753332220311896 MainEli SahirohNo ratings yet
- Circadian Rhythm Is A Biological Clock in Our Bodies That Synchronize With The 24-Hour Cycle of Day and NightDocument5 pagesCircadian Rhythm Is A Biological Clock in Our Bodies That Synchronize With The 24-Hour Cycle of Day and NightCherry LwinNo ratings yet
- TOP Science Grade 5 Worksheets Chapter 1Document11 pagesTOP Science Grade 5 Worksheets Chapter 1agustina simorangkirNo ratings yet
- Science G5: Quarter 2Document40 pagesScience G5: Quarter 2John ChristianNo ratings yet
- Pengantar FarmakologiDocument18 pagesPengantar FarmakologiHaidar AdiNo ratings yet
- Scientific CVDocument4 pagesScientific CVabilasha BalakrishnanNo ratings yet
- Jurassic Park TestDocument3 pagesJurassic Park TestІра ГусакNo ratings yet
- Human-Directed Dog AggressionDocument184 pagesHuman-Directed Dog AggressionTanya Vane WayNo ratings yet
- Zoo Lab Midterm ReviewerDocument8 pagesZoo Lab Midterm Reviewerandrew CruzNo ratings yet
- The Effects of Microwave Radiation On Microbial CuDocument8 pagesThe Effects of Microwave Radiation On Microbial CuFabien JaverliatNo ratings yet
- Mutations and Disorders worksheet-ANSDocument4 pagesMutations and Disorders worksheet-ANSGrace TaborNo ratings yet
- 19 - 28Document2 pages19 - 28Allen SoraNo ratings yet
- Significance and Implications of Mitosis and MeiosisDocument22 pagesSignificance and Implications of Mitosis and MeiosisAnilo James LedesmaNo ratings yet
- Biology The Dynamic Science 3rd Edition Russell Test Bank DownloadDocument21 pagesBiology The Dynamic Science 3rd Edition Russell Test Bank DownloadJamie Hayes100% (21)
- Chapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsDocument19 pagesChapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsNico Loko100% (1)
- Binder & Hibbett 2006 PDFDocument11 pagesBinder & Hibbett 2006 PDFSoto Agudelo RicardoNo ratings yet
- Gat B SyllabusDocument3 pagesGat B SyllabusDIKCHHA AGRAWALNo ratings yet
- Biology Chapter 3Document17 pagesBiology Chapter 3Fatima ZulqarnainNo ratings yet
- Learning Modules For General Biology 2Document12 pagesLearning Modules For General Biology 2Albert Rosete0% (1)
- Osteopetrosis - PediatricsDocument3 pagesOsteopetrosis - PediatricstaufiqtopNo ratings yet
- Dragon Genetics Lab-Part 1-Remote UlvDocument3 pagesDragon Genetics Lab-Part 1-Remote UlvstellaNo ratings yet
- Glycine & Serine NUB (IHB)Document4 pagesGlycine & Serine NUB (IHB)PIH SHTNo ratings yet