You might also like
- Imprinting DisordersDocument19 pagesImprinting Disorderssenuberh13No ratings yet
- Epigenetics in Reproductive MedicineDocument7 pagesEpigenetics in Reproductive MedicineVlad VochituNo ratings yet
- Lecture 10 Imprinting GeneDocument14 pagesLecture 10 Imprinting Genekamaran.tahaNo ratings yet
- Kotlyar 2020Document16 pagesKotlyar 2020Merab KvintradzeNo ratings yet
- (1479683X - European Journal of Endocrinology) Parental Genomic Imprinting in EndocrinopathiesDocument9 pages(1479683X - European Journal of Endocrinology) Parental Genomic Imprinting in EndocrinopathiesRavia SharmaNo ratings yet
- Im ProntaDocument12 pagesIm ProntaArturo MunozNo ratings yet
- PWS-Obesity Due To Genomic ImprintingDocument12 pagesPWS-Obesity Due To Genomic ImprintingTere HirokoNo ratings yet
- Developmental Genetics and Pharmacogeneticsdoc4301Document8 pagesDevelopmental Genetics and Pharmacogeneticsdoc4301sayednourNo ratings yet
- Genetic Basis of Oral DiseasesDocument23 pagesGenetic Basis of Oral Diseasesshanima sherinNo ratings yet
- Transgenerational Epigenetic Inheritance in Mammals: How Good Is The Evidence?Document9 pagesTransgenerational Epigenetic Inheritance in Mammals: How Good Is The Evidence?Juan Guillermo BuenaventuraNo ratings yet
- Chapter 8 - The Human Epigenome Implications For The Unde - 2009 - Molecular PaDocument13 pagesChapter 8 - The Human Epigenome Implications For The Unde - 2009 - Molecular PaSELNo ratings yet
- Genetics in OrthodonticsDocument56 pagesGenetics in OrthodonticsSadhu AbhijeetNo ratings yet
- Backgrounder: Epigenetics and Imprinted Genes: Jdowner1@jhmi - EduDocument4 pagesBackgrounder: Epigenetics and Imprinted Genes: Jdowner1@jhmi - Eduaryaa2020No ratings yet
- Seminar Presentation On GeneticsDocument60 pagesSeminar Presentation On GeneticsHardeep KaurNo ratings yet
- 53 Hamada Et Al Genetics and Male InfertilityDocument45 pages53 Hamada Et Al Genetics and Male InfertilitynehaNo ratings yet
- SUMMARYDocument2 pagesSUMMARYKathleen Laum CabanlitNo ratings yet
- Molecular and Genetic Aspects of AgingDocument6 pagesMolecular and Genetic Aspects of AgingRahul surya BobbiliNo ratings yet
- Genetic DisorderDocument13 pagesGenetic DisorderLucreatia RynjahNo ratings yet
- Genetic Basis of DiseaseDocument6 pagesGenetic Basis of Diseasedr saadia anjumNo ratings yet
- Genes, Environment and Inheritance - MG 4Document30 pagesGenes, Environment and Inheritance - MG 4snowrose2609No ratings yet
- Oncogenes and Tumour Suppressor GenesDocument59 pagesOncogenes and Tumour Suppressor GenesXutjie FairdianaNo ratings yet
- Pediatric Genetics Dr. Samed AlsalmiDocument166 pagesPediatric Genetics Dr. Samed AlsalmiabhivnairNo ratings yet
- Genetic Factors of Childhood CancerDocument3 pagesGenetic Factors of Childhood CancerDima HarounNo ratings yet
- PHD Credit Seminar 2 Punjab Agricultural University, LudhianaDocument141 pagesPHD Credit Seminar 2 Punjab Agricultural University, LudhianaSUSHANT DHARNo ratings yet
- Role of Genetics in Orthodontics - Simplified ConceptDocument5 pagesRole of Genetics in Orthodontics - Simplified ConceptInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Genetics and Orthodontics-FinalDocument96 pagesGenetics and Orthodontics-FinalRaj SinghNo ratings yet
- Lectu RaDocument5 pagesLectu RaMariaNo ratings yet
- 2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease MechanismsDocument12 pages2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease Mechanismsmarej312No ratings yet
- Precision Medicine NOTESDocument50 pagesPrecision Medicine NOTESNishtha KhannaNo ratings yet
- JojoDocument3 pagesJojoHui Robert ChengNo ratings yet
- Bio 1Document3 pagesBio 1James TolentinoNo ratings yet
- Onco Ch23 ESFTDocument31 pagesOnco Ch23 ESFTIlincaNo ratings yet
- Harrison's Principle of Internal Medicine 19thDocument113 pagesHarrison's Principle of Internal Medicine 19thTuan NguyenNo ratings yet
- Congenital Anomalies: Dr. Gara Samara B., SE, MMR, M.SC., M.Si - MedDocument59 pagesCongenital Anomalies: Dr. Gara Samara B., SE, MMR, M.SC., M.Si - MediikNo ratings yet
- Genetic Approach ToDocument15 pagesGenetic Approach TovipinNo ratings yet
- GEN3051 Lecture 1: Human Genes and Human Genetic DisordersDocument5 pagesGEN3051 Lecture 1: Human Genes and Human Genetic DisordersAlessander Leyendecker JuniorNo ratings yet
- Neoplasia: in These Lectures You Will Understand The FollowingDocument23 pagesNeoplasia: in These Lectures You Will Understand The FollowingpriyaNo ratings yet
- Genes 14 00585 PDFDocument13 pagesGenes 14 00585 PDFandy sotoNo ratings yet
- What Is The EpigenomeDocument2 pagesWhat Is The EpigenomeWA ODE DWI DANINGRAT 1306350371No ratings yet
- Dimorfismo SexualDocument21 pagesDimorfismo SexualDiana CortesNo ratings yet
- Pisharody Genetic CounselingDocument15 pagesPisharody Genetic Counselingapi-228136529No ratings yet
- Y Chromosome InfertilityDocument4 pagesY Chromosome Infertilityguido guida acevedoNo ratings yet
- Ameloblastoma CasesDocument52 pagesAmeloblastoma Casesdr arjunNo ratings yet
- 4 - 1 - Introduction of GenomeDocument13 pages4 - 1 - Introduction of GenomeALNAKINo ratings yet
- Garvan Institute of Medical Research - Genetics RefreshnerDocument17 pagesGarvan Institute of Medical Research - Genetics RefreshnerLuisaReyesNo ratings yet
- 2-Gen-Sept-14-Inheritance of Complex Disorders 2021 Post AheadDocument61 pages2-Gen-Sept-14-Inheritance of Complex Disorders 2021 Post Aheadn-gorNo ratings yet
- Can Environmental Factors Acting On An Organism Cause Inherited Changes in Phenotype in Its Descendants?Document3 pagesCan Environmental Factors Acting On An Organism Cause Inherited Changes in Phenotype in Its Descendants?DanNo ratings yet
- Medical Gen 1 TopicDocument28 pagesMedical Gen 1 TopicharshitaNo ratings yet
- X Extra-NuclearDocument11 pagesX Extra-Nuclearnsjunnarkar100% (1)
- Framework For Maternal and Child Health NursingDocument147 pagesFramework For Maternal and Child Health NursingRouie Björn ABrianNo ratings yet
- Misconceptions About The Use of Genetic Tests in PopulationsDocument4 pagesMisconceptions About The Use of Genetic Tests in PopulationsAneeshNo ratings yet
- Pathology AssignmentDocument61 pagesPathology AssignmentKepa KepaNo ratings yet
- Genetic TestingDocument18 pagesGenetic Testingneha100% (1)
- Overgrowth Syndromes: Andrew C. Edmondson Jennifer M. KalishDocument8 pagesOvergrowth Syndromes: Andrew C. Edmondson Jennifer M. KalishJuan Antonio Herrera LealNo ratings yet
- Genodermatoses: General ConsiderationsDocument21 pagesGenodermatoses: General ConsiderationsCyd Buenaflor-MadayagNo ratings yet
- Beckwith-Wiedemann SyndromeDocument6 pagesBeckwith-Wiedemann SyndromeAhmad Althof Malihul AdiNo ratings yet
- 13 GMppt-chapter16-2019 (Ok)Document17 pages13 GMppt-chapter16-2019 (Ok)Kw ChanNo ratings yet
- Louise Wilkins-Haug, MD, PHD Deborah Levine, MD Helen V Firth, DM, FRCP, DCH Vanessa A Barss, MD, Facog Contributor Disclosures Peer Review ProcessDocument10 pagesLouise Wilkins-Haug, MD, PHD Deborah Levine, MD Helen V Firth, DM, FRCP, DCH Vanessa A Barss, MD, Facog Contributor Disclosures Peer Review ProcessArienne_Mae_A__6554No ratings yet
- 10th Cytology LectureDocument57 pages10th Cytology LectureRC SILVESTRENo ratings yet
- Biology Infertility InvestigatoryDocument42 pagesBiology Infertility InvestigatorykeerthivasanNo ratings yet
- Wubtaye Hailu ProposalDocument38 pagesWubtaye Hailu ProposalWub-taye HailuNo ratings yet
- Tay-Sachs Disease I. What Is Tay-Sachs Disease?Document2 pagesTay-Sachs Disease I. What Is Tay-Sachs Disease?DBA MurialdoNo ratings yet
- Abortion EVICTIONISM - IS - LIBERTARIAN - DEPARTURISM - IS - NOT - CRITDocument16 pagesAbortion EVICTIONISM - IS - LIBERTARIAN - DEPARTURISM - IS - NOT - CRITNolbert BriceñoNo ratings yet
- Plant Organ Systems and Their Functions Page 2Document6 pagesPlant Organ Systems and Their Functions Page 2Reigne RamirezNo ratings yet
- The Importance of Breast MilkDocument2 pagesThe Importance of Breast MilkDenver SignabenNo ratings yet
- Obstetric Neurological Injuries: ReviewDocument8 pagesObstetric Neurological Injuries: Reviewaish25No ratings yet
- Immature Schistosoma ReflexDocument2 pagesImmature Schistosoma Reflexpramod kumarNo ratings yet
- Fetal Heart Rate Monitoring CompressedDocument279 pagesFetal Heart Rate Monitoring Compresseddr. syah sembung wasiso100% (1)
- Anatomy & Physiology - Male Reproductive SystemDocument2 pagesAnatomy & Physiology - Male Reproductive Systemkristel ludangcoNo ratings yet
- A Study To Assess The Effectiveness of Structured Teaching Program On Preconception Care PCC Among Women Studying at Selected College, BangaloreDocument4 pagesA Study To Assess The Effectiveness of Structured Teaching Program On Preconception Care PCC Among Women Studying at Selected College, BangaloreResearch ParkNo ratings yet
- Organizational Structure BnapDocument15 pagesOrganizational Structure BnapHenry Kahal Orio Jr.No ratings yet
- Emergency Contraception Knowledge, Attitude and Practices Among Female College Students atDocument14 pagesEmergency Contraception Knowledge, Attitude and Practices Among Female College Students atKIU PUBLICATION AND EXTENSIONNo ratings yet
- Judicial AND Xtra Udicial Onfessions: Health LawDocument27 pagesJudicial AND Xtra Udicial Onfessions: Health LawPranavNo ratings yet
- Agumentative EssayDocument3 pagesAgumentative EssaySherre Nicole CuentaNo ratings yet
- Intervensi DomainDocument92 pagesIntervensi DomainSylvia DamayantiNo ratings yet
- Livro 1 - 003 - Abortion Leading Cause of Death in 2018 With 41 Million KilledDocument3 pagesLivro 1 - 003 - Abortion Leading Cause of Death in 2018 With 41 Million KilledLeonardo OliveiraNo ratings yet
- AUBF Amniotic FluidDocument6 pagesAUBF Amniotic FluidRichelyn Grace B. VenusNo ratings yet
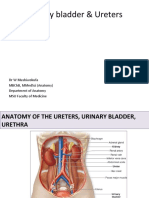
- Urinary Bladder & Ureters: DR W Mushiwokufa MBCHB, Mmedsci (Anatomy) Department of Anatomy Msu Faculty of MedicineDocument27 pagesUrinary Bladder & Ureters: DR W Mushiwokufa MBCHB, Mmedsci (Anatomy) Department of Anatomy Msu Faculty of MedicineLeona Tinotenda ManjeruNo ratings yet
- Antenatal AssessmentDocument67 pagesAntenatal Assessmenthiral mistry100% (2)
- Family Planning/contraception Methods: Key FactsDocument4 pagesFamily Planning/contraception Methods: Key FactsCheesy YoguNo ratings yet
- CSEC Biology June 2021 P2Document21 pagesCSEC Biology June 2021 P2Rianna SamsinghNo ratings yet
- Access To Maternity Healthcare Service in Rural Bangladesh. A Study Area Sariakandi Upazila of Bogura DistrictDocument13 pagesAccess To Maternity Healthcare Service in Rural Bangladesh. A Study Area Sariakandi Upazila of Bogura DistrictSajibNo ratings yet
- Pediatric Nursing 1الجزء الأولDocument291 pagesPediatric Nursing 1الجزء الأولVin Lorenzo CampbellNo ratings yet
- Aogs 12055Document8 pagesAogs 12055Wendy ThamrinNo ratings yet
- Tugas Cause and Effect Kelas ADocument1 pageTugas Cause and Effect Kelas Amazidah xiuNo ratings yet
- Stud Service ContractDocument3 pagesStud Service ContractTaccad ReydennNo ratings yet
- Nutrition Through The Life Cycle 6th Edition Brown Test Bank 1Document22 pagesNutrition Through The Life Cycle 6th Edition Brown Test Bank 1matthew100% (55)
- Prometric 2 PDFDocument24 pagesPrometric 2 PDFKatherine Joy ApquisNo ratings yet
- Agony AuntDocument2 pagesAgony AuntMariaPatriciaPaolantonioNo ratings yet