You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Questions About Behavioral Function (Qabf)Document3 pagesQuestions About Behavioral Function (Qabf)Registro psiNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Cognitive Training and Cognitive Rehabilitation For People With Early-Stage Alzheimer's Disease - Review - Clare & WoodsDocument18 pagesCognitive Training and Cognitive Rehabilitation For People With Early-Stage Alzheimer's Disease - Review - Clare & WoodsRegistro psiNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Role of Personality and Coping Style in Relation To Awareness Seiffer & Clare 2005Document8 pagesThe Role of Personality and Coping Style in Relation To Awareness Seiffer & Clare 2005Registro psiNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Measure Report Child CBCLDocument7 pagesMeasure Report Child CBCLEliza DumitruNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Knowing Where and Knowinh Mhat A Double Dissociation Barbara Wilson & Linda Clare 1997Document13 pagesKnowing Where and Knowinh Mhat A Double Dissociation Barbara Wilson & Linda Clare 1997Registro psiNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Awarenes in Dementia by Bob Woods 2005Document8 pagesAwarenes in Dementia by Bob Woods 2005Registro psiNo ratings yet
- Are Faces Special in AD Linda Clare 2007Document9 pagesAre Faces Special in AD Linda Clare 2007Registro psiNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Prevalence and Recognition of Major Depression Among Low Level Aged Care Residents PDFDocument8 pagesThe Prevalence and Recognition of Major Depression Among Low Level Aged Care Residents PDFRegistro psiNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Cognitive RehabilitationDocument10 pagesCognitive RehabilitationGloria ÁlvarezNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Topgraphical DisorientationDocument21 pagesTopgraphical DisorientationRegistro psiNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- O Que É Reserva Cognitiva PDFDocument13 pagesO Que É Reserva Cognitiva PDFBrenda BorgesNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Where Do PerseverationsDocument11 pagesWhere Do PerseverationsRegistro psiNo ratings yet
- Cognitive Stimulation and DementiaDocument5 pagesCognitive Stimulation and DementiaRegistro psiNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- TheprogressionofcognitionDocument7 pagesTheprogressionofcognitionRegistro psiNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Belief in A Just WorldDocument7 pagesThe Belief in A Just WorldRegistro psiNo ratings yet
- AcommonneuralnetworkDocument9 pagesAcommonneuralnetworkRegistro psiNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Theses Theses Theses Theses Theses: Benito Pereira DamascenoDocument3 pagesTheses Theses Theses Theses Theses: Benito Pereira DamascenoRegistro psiNo ratings yet
- Bases Neurobiologicas de Las Dificultades de AprendizajeDocument5 pagesBases Neurobiologicas de Las Dificultades de Aprendizajeapi-3824526100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Verbal Fluency Predicts Mortality in Alzheimer DiseaseDocument7 pagesVerbal Fluency Predicts Mortality in Alzheimer DiseaseRegistro psiNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Boston Naming Test: Performance of Brazilian Population From São PauloDocument8 pagesBoston Naming Test: Performance of Brazilian Population From São PauloRegistro psiNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Meanings Older People GiveDocument18 pagesThe Meanings Older People GiveRegistro psiNo ratings yet
- Aval. Mem Ria - Alzheimer PDFDocument8 pagesAval. Mem Ria - Alzheimer PDFRegistro psiNo ratings yet
- Book Review PDFDocument4 pagesBook Review PDFRegistro psiNo ratings yet
- Categorical and Coordinate Spatial Representations 2008Document8 pagesCategorical and Coordinate Spatial Representations 2008Registro psiNo ratings yet
- Action Errors and Dressing Disability After Stroke PDFDocument19 pagesAction Errors and Dressing Disability After Stroke PDFRegistro psiNo ratings yet
- An Investigation of The Differential Effect of Self GenerationDocument21 pagesAn Investigation of The Differential Effect of Self GenerationRegistro psiNo ratings yet
- Prospective Memory in Adults With Traumatic Brain InjuryDocument22 pagesProspective Memory in Adults With Traumatic Brain InjuryRegistro psiNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Topgraphical DisorientationDocument21 pagesTopgraphical DisorientationRegistro psiNo ratings yet
- The Sensitivity and Specificity of The Middlesex Elderly Assessment of Mental StateDocument14 pagesThe Sensitivity and Specificity of The Middlesex Elderly Assessment of Mental StateRegistro psiNo ratings yet
- Spatial Memory and The Human Hippocampus 2007Document6 pagesSpatial Memory and The Human Hippocampus 2007Registro psiNo ratings yet
- Pharm-Immuno 9-11 Cell Mediated Immunity: Dr. Saber HusseinDocument48 pagesPharm-Immuno 9-11 Cell Mediated Immunity: Dr. Saber Husseinmmoney1No ratings yet
- Cerebrospinal Fluid Flow - Anatomy and Functions - KenhubDocument8 pagesCerebrospinal Fluid Flow - Anatomy and Functions - KenhubSajal SahaNo ratings yet
- Fetal Circulation ORIGINALDocument13 pagesFetal Circulation ORIGINALombir007No ratings yet
- Autoimmune Disorders - AgungDocument40 pagesAutoimmune Disorders - AgungalgutNo ratings yet
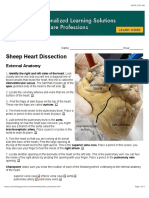
- Sheep Heart DissectionDocument3 pagesSheep Heart DissectionMorgan DanielsNo ratings yet
- Physiological Changes in PregnancyDocument18 pagesPhysiological Changes in Pregnancysubvig100% (3)
- Iycf AsiaDocument68 pagesIycf AsiaYahye CMNo ratings yet
- Anatomy & Physiology Muscular SystemDocument14 pagesAnatomy & Physiology Muscular SystemYOPO KITZ ANGELONo ratings yet
- Thyrotoxic Crisis ( Thyroid Storm')Document7 pagesThyrotoxic Crisis ( Thyroid Storm')Roopa Lexmy MuniandyNo ratings yet
- Clone 123C3: Monoclonal Mouse Anti-Human CD56 Code M7304Document3 pagesClone 123C3: Monoclonal Mouse Anti-Human CD56 Code M7304Jaimier CajandabNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Physiology of Breast Milk ProductionDocument17 pagesPhysiology of Breast Milk ProductionOlivia TegliaNo ratings yet
- Motor System: G. Arveladze N. MalashkhiaDocument17 pagesMotor System: G. Arveladze N. MalashkhialuckyNo ratings yet
- Artikel Andika RediputraDocument6 pagesArtikel Andika RediputraproluvieslacusNo ratings yet
- Endocrine System - outlINEDocument4 pagesEndocrine System - outlINERalph NilloNo ratings yet
- Organ Donation: A Life-Saving GiftDocument3 pagesOrgan Donation: A Life-Saving GiftGideon CropNo ratings yet
- Perdev ReportingDocument30 pagesPerdev ReportingIgrid Kyla LayesaNo ratings yet
- Ultimate Breastfeeding Guide The Thompson MethodDocument48 pagesUltimate Breastfeeding Guide The Thompson MethodlisibulavouNo ratings yet
- Duffau (2018) - Error de BrocaDocument9 pagesDuffau (2018) - Error de BrocaMaría Cordero Varela0% (1)
- 1.1 The Human Body An OrientationDocument59 pages1.1 The Human Body An OrientationDon KeyNo ratings yet
- Duction Trophy A + Trophy Algesic A + AlgosDocument15 pagesDuction Trophy A + Trophy Algesic A + AlgosEmad AlryashiNo ratings yet
- FULL Download Ebook PDF Immunology A Short Course 7th Edition PDF EbookDocument41 pagesFULL Download Ebook PDF Immunology A Short Course 7th Edition PDF Ebookangel.lain349100% (35)
- Respiratory TransesDocument4 pagesRespiratory TransesAriane Shane BidoNo ratings yet
- Benign Breast DiseasesDocument35 pagesBenign Breast DiseasesBhim Nath KharelNo ratings yet
- 511 FullDocument7 pages511 FullMarcela MurafaNo ratings yet
- DR - Elamaran.E Senior Resident Dept. of CTVS, JIPMERDocument43 pagesDR - Elamaran.E Senior Resident Dept. of CTVS, JIPMEREla MaranNo ratings yet
- Adrenal CortexDocument8 pagesAdrenal CortexEdward XiamNo ratings yet
- Baroreceptor Reflex (Costanzo 3rd Ed)Document2 pagesBaroreceptor Reflex (Costanzo 3rd Ed)Gita FebrianyNo ratings yet
- Digestive System of A Typical Fish (Rohu)Document13 pagesDigestive System of A Typical Fish (Rohu)Ashok AdhikariNo ratings yet
- Thyroid Healthy Iodine, Lose Weight, Look Beautiful and Live The Life You Imagine (Suzy Cohen RPH, David Brownstein) (Z-Library)Document308 pagesThyroid Healthy Iodine, Lose Weight, Look Beautiful and Live The Life You Imagine (Suzy Cohen RPH, David Brownstein) (Z-Library)Анна Варданян100% (1)
- Glandular AffectionsDocument3 pagesGlandular AffectionsRajiv Ranjan SinhaNo ratings yet