You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Cardioprotection of Vitexin On Myocardial IschemiaDocument17 pagesCardioprotection of Vitexin On Myocardial IschemiaVampyrgirlNo ratings yet
- Antidepressant Like Effects and Mechanisms of Flavanoids and Related AnaloguesDocument11 pagesAntidepressant Like Effects and Mechanisms of Flavanoids and Related AnaloguesVampyrgirlNo ratings yet
- Antidepressant-Like Effects of Melatonin in The Mouse Chronic Mild Stress ModelDocument5 pagesAntidepressant-Like Effects of Melatonin in The Mouse Chronic Mild Stress ModelVampyrgirlNo ratings yet
- Assessing Mouse Behaviour Throughout The Light-Dark Cycle Using Automated In-Cage Analysis ToolsDocument11 pagesAssessing Mouse Behaviour Throughout The Light-Dark Cycle Using Automated In-Cage Analysis ToolsVampyrgirlNo ratings yet
- An Efficient Chronic Unpredictable Stress Protocol To Induce Stress-Related Responses in C57BL-6 MiceDocument11 pagesAn Efficient Chronic Unpredictable Stress Protocol To Induce Stress-Related Responses in C57BL-6 MiceVampyrgirlNo ratings yet
- An Unpredictable Chronic Mild Stress Protocol For Instigating Depressive Symptoms Behavioral Changes and Negative Health Outcomes in RodentsDocument8 pagesAn Unpredictable Chronic Mild Stress Protocol For Instigating Depressive Symptoms Behavioral Changes and Negative Health Outcomes in RodentsVampyrgirlNo ratings yet
- Altered Aortic Vascular Reactivity in The Unpredictable Chronic Mild Stress Model of Depression in Mice UCMS Causes Relaxation Impairment To AChDocument7 pagesAltered Aortic Vascular Reactivity in The Unpredictable Chronic Mild Stress Model of Depression in Mice UCMS Causes Relaxation Impairment To AChVampyrgirlNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Microbiology Principles and Explorations 10th Edition Black Test BankDocument36 pagesMicrobiology Principles and Explorations 10th Edition Black Test Bankdavidnelsonseyjdmpoaz100% (12)
- Optic Nerve MeningiomasDocument1 pageOptic Nerve MeningiomasMohammadAwitNo ratings yet
- ArticuloDocument11 pagesArticuloalvarogascareinaNo ratings yet
- Pathology Lecture 7 - LiverDocument11 pagesPathology Lecture 7 - Livercgao30No ratings yet
- Introduction to Laboratory ManagementDocument7 pagesIntroduction to Laboratory ManagementBea Reen BurgosNo ratings yet
- Nursing Care of The Client With CancerDocument15 pagesNursing Care of The Client With CancerAlvin M Alcayno0% (1)
- HCC HarrisonDocument5 pagesHCC HarrisonJesly CharliesNo ratings yet
- UNIT 2 - Micro-Organisms in Food NDocument29 pagesUNIT 2 - Micro-Organisms in Food NShiv YadavNo ratings yet
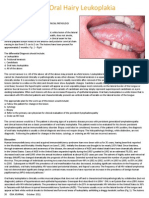
- Hairy LeukoplakiaDocument2 pagesHairy LeukoplakiaEka Septariana PuspaNo ratings yet
- Pharmaceutical Biotechnology: Tushar Kapur Ranbir Negi Samarth Agarwal Prakhar DubeyDocument16 pagesPharmaceutical Biotechnology: Tushar Kapur Ranbir Negi Samarth Agarwal Prakhar DubeyRanbir100% (1)
- Interference Testing: Clin Biochem Rev. 2008 August 29 (Supplement (I) ) : S43-S48. PMCID: PMC2556582Document9 pagesInterference Testing: Clin Biochem Rev. 2008 August 29 (Supplement (I) ) : S43-S48. PMCID: PMC2556582Kesatria Bayu KencanaNo ratings yet
- Feedback Control MechanismsDocument4 pagesFeedback Control MechanismsJividan SasidranNo ratings yet
- Craig Venter Mycoplasma LaboratoriumDocument24 pagesCraig Venter Mycoplasma LaboratoriumHaveit12No ratings yet
- Gram Staining PDFDocument6 pagesGram Staining PDFMaria Chacón CarbajalNo ratings yet
- Microbiota Intestinal e LúpusDocument8 pagesMicrobiota Intestinal e Lúpusadriana félixNo ratings yet
- Forensic ScienceDocument10 pagesForensic ScienceVyomaNo ratings yet
- Chatterjee Et Al., 2017 - IGEDocument5 pagesChatterjee Et Al., 2017 - IGEPabitra SahaNo ratings yet
- RBC MembraneDocument4 pagesRBC MembraneJustine Claire M. NamocatcatNo ratings yet
- ICD-10 codes for mental disordersDocument5 pagesICD-10 codes for mental disordersAsri ParantriNo ratings yet
- Theories of Ageing: Presented by DR - Pratyusha KaligotlaDocument13 pagesTheories of Ageing: Presented by DR - Pratyusha KaligotlaSHAIK SHABEENANo ratings yet
- Neurotransmitter, Peptide and Cytokine Processes in Relation to Depressive Disorder and Neurodegenerative ComorbidityDocument74 pagesNeurotransmitter, Peptide and Cytokine Processes in Relation to Depressive Disorder and Neurodegenerative ComorbidityShawn HayleyNo ratings yet
- AXP Ii System: Cordlife IntroducesDocument2 pagesAXP Ii System: Cordlife Introducessanath kumarNo ratings yet
- Development of The Nervous SystemDocument5 pagesDevelopment of The Nervous SystemChaGonzalesNo ratings yet
- Kami Export - 9.4 - UsingTheHardyWeinbergEquation - Worksheet Danielle Ngambia TchetmiDocument3 pagesKami Export - 9.4 - UsingTheHardyWeinbergEquation - Worksheet Danielle Ngambia TchetmijongambiaNo ratings yet
- Cardiac Enzymes and Markers For Myocardial InfarctionDocument11 pagesCardiac Enzymes and Markers For Myocardial InfarctionFitria NingsihhNo ratings yet
- Test Bank For Principles of Animal Physiology 2nd Edition Christopher D MoyesDocument303 pagesTest Bank For Principles of Animal Physiology 2nd Edition Christopher D MoyesKathy Galloway100% (26)
- Types of Specimen Collection ContainersDocument7 pagesTypes of Specimen Collection ContainersLucas Dei AmengorNo ratings yet
- TSEBTDocument17 pagesTSEBTcornejo1No ratings yet
- Book of Apstracts 6th Congress SGSDocument160 pagesBook of Apstracts 6th Congress SGSmilica cucuzNo ratings yet
- Hamza DissertDocument108 pagesHamza DissertMobin Ur Rehman KhanNo ratings yet