You might also like
- Social Anxiety and The Severity and Typography of Stuttering in AdolescentsDocument14 pagesSocial Anxiety and The Severity and Typography of Stuttering in Adolescentsszabin.gereNo ratings yet
- Fear of Negative Evaluation and The Hypervigilance-Avoidance Hypothesis: An Eye-Tracking StudyDocument7 pagesFear of Negative Evaluation and The Hypervigilance-Avoidance Hypothesis: An Eye-Tracking Studyselamet apriyantoNo ratings yet
- nst025 PDFDocument9 pagesnst025 PDFRavennaNo ratings yet
- Frontal Late Positivity in Dental Phobia: A Study On Gender DifferencesDocument7 pagesFrontal Late Positivity in Dental Phobia: A Study On Gender DifferencesSung Soon ChangNo ratings yet
- Vriends Et Al. (2012) - Associative Learning in Flying PhobiaDocument6 pagesVriends Et Al. (2012) - Associative Learning in Flying PhobiaEcker HernándezNo ratings yet
- Neural Correlates of Attachment Trauma in Borderline Personality Disorder: A Functional Magnetic Resonance Imaging StudyDocument13 pagesNeural Correlates of Attachment Trauma in Borderline Personality Disorder: A Functional Magnetic Resonance Imaging StudyAlina AlexandrescuNo ratings yet
- Individual Differences in Childrenõs Performance During An Emotional Stroop Task: A Behavioral and Electrophysiological StudyDocument19 pagesIndividual Differences in Childrenõs Performance During An Emotional Stroop Task: A Behavioral and Electrophysiological StudyAdriana VeronicaNo ratings yet
- Empathy and Symptoms DimensionsDocument9 pagesEmpathy and Symptoms DimensionsReem FzNo ratings yet
- 2009 - Intolerance of Uncertainty and Social Anxiety. - Boelen, Reijntjes PDFDocument6 pages2009 - Intolerance of Uncertainty and Social Anxiety. - Boelen, Reijntjes PDFFabián MaeroNo ratings yet
- Neural Bases of Antisocial Behavior: A Voxel-Based Meta-AnalysisDocument9 pagesNeural Bases of Antisocial Behavior: A Voxel-Based Meta-AnalysisYeremias EdwinNo ratings yet
- Emotional Intelligence and Psychological Resili 2011 Personality and IndividDocument6 pagesEmotional Intelligence and Psychological Resili 2011 Personality and IndividTorburam LhamNo ratings yet
- AnxStut - JFD - 2004 - Social Anxiety in Stuttering - Measuring Negative Social ExpectanciesDocument12 pagesAnxStut - JFD - 2004 - Social Anxiety in Stuttering - Measuring Negative Social ExpectanciesfpNo ratings yet
- Vitoratou Et Al 2020. Selective Sound Sensitivity Syndrome Scale S5.Document47 pagesVitoratou Et Al 2020. Selective Sound Sensitivity Syndrome Scale S5.Júlia Petúlia SolNo ratings yet
- Theory of mind and cognitive flexibility in schizophreniaDocument9 pagesTheory of mind and cognitive flexibility in schizophreniaMichuuu AguilarNo ratings yet
- Fnbeh 08 00241 PDFDocument14 pagesFnbeh 08 00241 PDFymintriagoNo ratings yet
- Rouw 2017Document27 pagesRouw 2017Francisco Urdiales ReinaNo ratings yet
- Dorsal - Ventral Parcellation of The Amygdala Relevance To Impulsivity and AggressionDocument7 pagesDorsal - Ventral Parcellation of The Amygdala Relevance To Impulsivity and AggressionfernandogfcnsNo ratings yet
- 1 s2.0 S0306453021000810 MainDocument9 pages1 s2.0 S0306453021000810 MainLuíza PintoNo ratings yet
- Appraisal Psychology, Neurobiology, and LanguageDocument20 pagesAppraisal Psychology, Neurobiology, and Languagemarjeybob99No ratings yet
- Effects of Worry and Progressive Relaxation On The Reduction of Fear in Speech Phobia: An Investigation of Situational ExposureDocument3 pagesEffects of Worry and Progressive Relaxation On The Reduction of Fear in Speech Phobia: An Investigation of Situational Exposurealmendras_amargasNo ratings yet
- Alex o Poulos 2018Document25 pagesAlex o Poulos 2018Sung Soon ChangNo ratings yet
- A Functional and Structural Study of Emotion and Face Processing in Children With AutismDocument10 pagesA Functional and Structural Study of Emotion and Face Processing in Children With AutismPadma Vasavi K NNo ratings yet
- Art 1 Dra. F PDFDocument8 pagesArt 1 Dra. F PDFNatalia Aristizábal DuqueNo ratings yet
- Psychopathy Aggression and Emotion ProceDocument5 pagesPsychopathy Aggression and Emotion Procetim radellNo ratings yet
- Teachman Et Al 2001 JAP SnakeSpiderIATDocument10 pagesTeachman Et Al 2001 JAP SnakeSpiderIATAiden P GreggNo ratings yet
- Neurobiological Underpinnings of Shame and Guilt: A Pilot fMRI StudyDocument8 pagesNeurobiological Underpinnings of Shame and Guilt: A Pilot fMRI StudyJose EscobarNo ratings yet
- Miu PsychRes 2012Document6 pagesMiu PsychRes 2012SumedreaCristianNo ratings yet
- The Relationship Between Anxiety and StuDocument14 pagesThe Relationship Between Anxiety and Stumajid mirzaeeNo ratings yet
- Soc Cogn Affect Neurosci-2009-Cri-an-399-408Document10 pagesSoc Cogn Affect Neurosci-2009-Cri-an-399-408LooTonyNo ratings yet
- Review Maltreatment&BrainDocument26 pagesReview Maltreatment&BrainMaudeNo ratings yet
- Annual Research Review Enduring Neurobiological Effects of Childhood Abuse and NeglectDocument26 pagesAnnual Research Review Enduring Neurobiological Effects of Childhood Abuse and NeglectDonald Cabrera AstudilloNo ratings yet
- Brain and Cognition: Anna Weinberg, E. David Klonsky, Greg HajcakDocument8 pagesBrain and Cognition: Anna Weinberg, E. David Klonsky, Greg HajcakGheorghe SecuNo ratings yet
- Brain and Cognition: Anna Weinberg, E. David Klonsky, Greg HajcakDocument8 pagesBrain and Cognition: Anna Weinberg, E. David Klonsky, Greg HajcakGheorghe SecuNo ratings yet
- Fear and Phobias 1Document7 pagesFear and Phobias 1api-395480978No ratings yet
- Neural Systems For Recognition of Emotinal ProsodyDocument29 pagesNeural Systems For Recognition of Emotinal ProsodyagneswindramNo ratings yet
- Emotional Intelligence and Psychological Resilience To Negative Life EventsDocument6 pagesEmotional Intelligence and Psychological Resilience To Negative Life EventsBogdan Hadarag100% (1)
- Nyctophobia From Imagined to Realistic Fears of the DarkDocument9 pagesNyctophobia From Imagined to Realistic Fears of the DarkJosé Carlos Buesso JrNo ratings yet
- Wilhelm 2001Document22 pagesWilhelm 2001AlejandraNo ratings yet
- Limitations of Cognitive Control On Emotional Distraction - Congruency in The Color Stroop Task Does Not Modulate The Emotional Stroop EffectDocument21 pagesLimitations of Cognitive Control On Emotional Distraction - Congruency in The Color Stroop Task Does Not Modulate The Emotional Stroop EffectASNo ratings yet
- vanhonk2011Document2 pagesvanhonk2011bonnie.a.omalleyNo ratings yet
- Exist. Anxiety in AdolescentsDocument9 pagesExist. Anxiety in Adolescentsluxlelito97No ratings yet
- Pineda 2009Document9 pagesPineda 2009yma ymiiNo ratings yet
- Simulation 102Document41 pagesSimulation 102nidhiNo ratings yet
- (2021) - The Relationship Between Alexithymia and Theory of Mind A Systematic ReviewDocument28 pages(2021) - The Relationship Between Alexithymia and Theory of Mind A Systematic ReviewBruno SallesNo ratings yet
- Entomology 1Document4 pagesEntomology 1Ozman NovelNo ratings yet
- Troiani Schultz 2013 Research AmygdalaDocument13 pagesTroiani Schultz 2013 Research AmygdalaElena SanduNo ratings yet
- The Relationships Between Stressful Life Events During Childhood and Differentiation of Self and Intergenerational Triangulation in AdulthoodDocument9 pagesThe Relationships Between Stressful Life Events During Childhood and Differentiation of Self and Intergenerational Triangulation in AdulthoodAna María González FNo ratings yet
- 1 s2.0 S0149763422003645 MainDocument18 pages1 s2.0 S0149763422003645 MainLilianne Mbengani LaranjeiraNo ratings yet
- Tâm LýDocument7 pagesTâm LýlinhvuyxinhNo ratings yet
- Car Thy 2009Document14 pagesCar Thy 2009AoiNo ratings yet
- Anger - Prosody CognitionDocument11 pagesAnger - Prosody Cognitionmarcu_ruxandraNo ratings yet
- Age of Onset of Dental AnxietyDocument7 pagesAge of Onset of Dental AnxietyHajar ChebliNo ratings yet
- Study 1 Music - MoodDocument9 pagesStudy 1 Music - MoodYomna ElmassryNo ratings yet
- Anxious SymptomsDocument7 pagesAnxious Symptomst mdeNo ratings yet
- Pechtel P11Document16 pagesPechtel P11cfrl25No ratings yet
- From Epiphenomenon To Biologically Important PhenomenaDocument4 pagesFrom Epiphenomenon To Biologically Important PhenomenaeierulloNo ratings yet
- The Unique and Conditional Effects of Interoceptive Exposure in TheDocument14 pagesThe Unique and Conditional Effects of Interoceptive Exposure in TheMARGARITA VICTORIA NINO SANDOVALNo ratings yet
- Contacto VisualDocument4 pagesContacto VisualSolange Medina AriasNo ratings yet
- Journal of Experimental Psychopathology: Justin W. Weeks, Peggy M. ZoccolaDocument23 pagesJournal of Experimental Psychopathology: Justin W. Weeks, Peggy M. Zoccolaselamet apriyantoNo ratings yet
- A Clinical Guide to the Treatment of the Human Stress ResponseFrom EverandA Clinical Guide to the Treatment of the Human Stress ResponseNo ratings yet
- Consolidated Guidelines OnDocument594 pagesConsolidated Guidelines OnRICHARD EDUARDO FERNANDEZ SAENZNo ratings yet
- LO SSS Week 5Document8 pagesLO SSS Week 5Tony StarkNo ratings yet
- Guideline Ascites 4UFb 2015Document96 pagesGuideline Ascites 4UFb 2015abrar aliNo ratings yet
- Guias ESC para Insuficiencia Cardiaca CronicaDocument128 pagesGuias ESC para Insuficiencia Cardiaca CronicaKarla HernandezNo ratings yet
- Guidelines On The Management of Ascites in CirrhosisDocument21 pagesGuidelines On The Management of Ascites in CirrhosisTony StarkNo ratings yet
- Guidelines For Intravenous Albumin Administration PDFDocument4 pagesGuidelines For Intravenous Albumin Administration PDFKunni MardhiyahNo ratings yet
- Beard 2011Document5 pagesBeard 2011Tony StarkNo ratings yet
- Groves 2016Document26 pagesGroves 2016Tony StarkNo ratings yet
- 2007 Ada DkaDocument9 pages2007 Ada DkaMetinee SutthivaiyakitNo ratings yet
- (Jojo's Bizarre Adventure) Giorno's Theme PDFDocument12 pages(Jojo's Bizarre Adventure) Giorno's Theme PDFFerrtasNo ratings yet
- 2006 Ryan PDFDocument17 pages2006 Ryan PDFDandara PalhanoNo ratings yet
- Accident Analysis and PreventionDocument8 pagesAccident Analysis and PreventionTony StarkNo ratings yet
- Summertime Cinnamons x Evening CinemaDocument5 pagesSummertime Cinnamons x Evening CinemaTony Stark100% (1)
- 10 3389@fpsyg 2018 00787Document15 pages10 3389@fpsyg 2018 00787Tony StarkNo ratings yet
- Rick and Morty - Evil Morty ThemeDocument2 pagesRick and Morty - Evil Morty ThemeTony StarkNo ratings yet
- 10 1155@2013@546342Document8 pages10 1155@2013@546342Tony StarkNo ratings yet
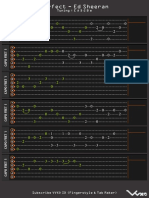
- Perfect TABDocument4 pagesPerfect TABMateus Gheorghiu100% (1)
- Coffee and Tea Consumption and Risk of Type 2 DiabetesDocument22 pagesCoffee and Tea Consumption and Risk of Type 2 DiabetesTony StarkNo ratings yet
- Giornos Theme PDFDocument2 pagesGiornos Theme PDFTony StarkNo ratings yet
- DPES Medical EthicsDocument17 pagesDPES Medical EthicsTony StarkNo ratings yet
- Grass Skirt Chase Ukulele Tabs SpongebobDocument2 pagesGrass Skirt Chase Ukulele Tabs SpongebobTony Stark100% (1)
- DPES Medical EthicsDocument17 pagesDPES Medical EthicsTony StarkNo ratings yet
- Food Impaction - Hirschfeld1930Document25 pagesFood Impaction - Hirschfeld1930Pockets Ain't EmptyNo ratings yet
- Chapter 11 Hormonal Coordination: 165 MinutesDocument60 pagesChapter 11 Hormonal Coordination: 165 MinutesAviBreezeNo ratings yet
- Dwnload Full Introduction To Radiologic and Imaging Sciences and Patient Care 6th Edition Adler Test Bank PDFDocument36 pagesDwnload Full Introduction To Radiologic and Imaging Sciences and Patient Care 6th Edition Adler Test Bank PDFChristopherNortonwqnmc100% (12)
- Guide Sports and Games - Around The WorldDocument29 pagesGuide Sports and Games - Around The WorldJoseph VEGANo ratings yet
- Insecticidal Activity of Tomato (Solanum Lycopersicum) Extract On American Cockroach (Periplenata Americana)Document7 pagesInsecticidal Activity of Tomato (Solanum Lycopersicum) Extract On American Cockroach (Periplenata Americana)heheeheheNo ratings yet
- Bodyweight Exercises For Extraordinary StrengthDocument129 pagesBodyweight Exercises For Extraordinary StrengthMattia Ferri86% (7)
- WLP-BLP GRADE 10 Week 8 L2-4 HDocument7 pagesWLP-BLP GRADE 10 Week 8 L2-4 HKalabit PengeNo ratings yet
- Situation of Drugs Use Among IDUsDocument52 pagesSituation of Drugs Use Among IDUsSuresh DhunganaNo ratings yet
- De Thi Tuyen Sinh Lop 10 THPT 20162017Document5 pagesDe Thi Tuyen Sinh Lop 10 THPT 20162017Huy HaNo ratings yet
- Mallari 397-414Document19 pagesMallari 397-414Trixia Louise NastorNo ratings yet
- 1.8 Annex 4 - BOPIS DCF FORM 2-BDocument2 pages1.8 Annex 4 - BOPIS DCF FORM 2-Bgulp_burp75% (4)
- Availability and Utilization of Medical Devices in Jimma Zone Hospitals, Southwest Ethiopia: A Case StudyDocument11 pagesAvailability and Utilization of Medical Devices in Jimma Zone Hospitals, Southwest Ethiopia: A Case StudyEzanaNo ratings yet
- Nursing Informatics in the Philippines History and IssuesDocument2 pagesNursing Informatics in the Philippines History and IssuesCorpus, Irene Zen P.No ratings yet
- The Long Term Effects of Cyberbullying On Adolescent Mental HealthDocument27 pagesThe Long Term Effects of Cyberbullying On Adolescent Mental Healthnelrose NavascaNo ratings yet
- Curative SARS-CoV-2 Test Results NegativeDocument2 pagesCurative SARS-CoV-2 Test Results Negativeluis torresNo ratings yet
- Interpersonal Problems and The Perception of Therapeutic Factors in Group TherapyDocument17 pagesInterpersonal Problems and The Perception of Therapeutic Factors in Group TherapyTasos TravasarosNo ratings yet
- Turn Table - LLT-750Document29 pagesTurn Table - LLT-750Sadashiva sahooNo ratings yet
- Multi Bandage Presentation - URIEL Meditex LTDDocument9 pagesMulti Bandage Presentation - URIEL Meditex LTDIsrael ExporterNo ratings yet
- 2.12 ICS Forms ICS 206Document3 pages2.12 ICS Forms ICS 206mdrrmo sinait100% (1)
- EssayDocument5 pagesEssayPranjal SrivastavaNo ratings yet
- Intellectual Disability and Assistive Technology: Opening The Gate WiderDocument4 pagesIntellectual Disability and Assistive Technology: Opening The Gate WiderBudi Darmawan DiswanNo ratings yet
- Nutrition Factor in Women in Backward AreasDocument7 pagesNutrition Factor in Women in Backward AreaspranjalNo ratings yet
- Lumi Juice CaseDocument6 pagesLumi Juice CaseIsmael BastidaNo ratings yet
- Writing - Task - 2-Chia Sẻ Tài Liệu Học IELTS 9.0Document301 pagesWriting - Task - 2-Chia Sẻ Tài Liệu Học IELTS 9.0Vũ Thái SơnNo ratings yet
- Certificate of Readiness To Enter Higher Surgical Training 2021Document5 pagesCertificate of Readiness To Enter Higher Surgical Training 2021mNo ratings yet
- Anti-Smoking Barangay ScorecardDocument2 pagesAnti-Smoking Barangay ScorecardBarangay PitaNo ratings yet
- Lesson Plan in Bread and Pastry Production Ncii I. ObjectivesDocument2 pagesLesson Plan in Bread and Pastry Production Ncii I. ObjectivesDezzelyn Balleta100% (1)
- Tissue CommitteeDocument3 pagesTissue Committeedjtudz100% (1)
- Climbing WorkoutsDocument59 pagesClimbing WorkoutsAlexandre Santos100% (8)
- Hazard Identification and Risk Assessment for Confined Space WorkDocument4 pagesHazard Identification and Risk Assessment for Confined Space Workaman anand100% (2)