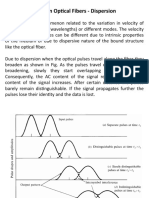
You might also like
- 2018 Identification of DNA Markers Related To Resistanc - PDF EngDocument9 pages2018 Identification of DNA Markers Related To Resistanc - PDF EngMay Htet AungNo ratings yet
- Mapping of Cotton-Melon Aphid Resistance in MelonDocument8 pagesMapping of Cotton-Melon Aphid Resistance in MelonanradeljaNo ratings yet
- Petzold-Maxwell 2013. Applying An Integrated Refuge To Manage Western Corn Rootworm Effects On Survival, Fitness, and Selection PressureDocument14 pagesPetzold-Maxwell 2013. Applying An Integrated Refuge To Manage Western Corn Rootworm Effects On Survival, Fitness, and Selection PressuresebastianNo ratings yet
- Colletotrichum Acutatum in Chili Pepper Capsicum Annuum L.) : Resistance To Anthracnose Caused by (Document4 pagesColletotrichum Acutatum in Chili Pepper Capsicum Annuum L.) : Resistance To Anthracnose Caused by (Pesol HaryantoNo ratings yet
- Inheritance of Resistance to Phytophthora Root Rot in Chili PepperDocument7 pagesInheritance of Resistance to Phytophthora Root Rot in Chili PepperChai Hong XuanNo ratings yet
- Cytological Effects of Blitox On Root Mitosis of Allium: Cepa LDocument7 pagesCytological Effects of Blitox On Root Mitosis of Allium: Cepa La lNo ratings yet
- Hypersensitive-Like Reaction Conferred by The Mex-1 Resistance Gene Against Meloidogyne Exigua in CoffeeDocument7 pagesHypersensitive-Like Reaction Conferred by The Mex-1 Resistance Gene Against Meloidogyne Exigua in CoffeeTrần Bảo ThắngNo ratings yet
- Mekanisme Ketahanan Kentang (Solanum Tuberosum) Terhadap Nematoda Sista Kuning (Globodera Rostochiensis)Document8 pagesMekanisme Ketahanan Kentang (Solanum Tuberosum) Terhadap Nematoda Sista Kuning (Globodera Rostochiensis)julpaidukNo ratings yet
- 30 36 PDFDocument7 pages30 36 PDFray m deraniaNo ratings yet
- Diversity in Virulence of Xanthomonas Oryzae Pv. OryzaeDocument11 pagesDiversity in Virulence of Xanthomonas Oryzae Pv. OryzaeSri MulyaniNo ratings yet
- Trangenic TrigoDocument11 pagesTrangenic TrigoCesarRamNo ratings yet
- Eivaziankary2018 PDFDocument8 pagesEivaziankary2018 PDFGiribabu PalaniappanNo ratings yet
- Dhungana Et Al-2015-Journal of Agronomy and Crop ScienceDocument7 pagesDhungana Et Al-2015-Journal of Agronomy and Crop ScienceAzhari RizalNo ratings yet
- Biocontrol PDFDocument6 pagesBiocontrol PDFWesh De GeminosNo ratings yet
- Temporal Dynamics of Potato Leaf Metabolites After Phytophthora Infestans InoculationDocument17 pagesTemporal Dynamics of Potato Leaf Metabolites After Phytophthora Infestans InoculationKeyla GonzálezNo ratings yet
- Breeding For Fusarium Wilt Resistance and Some Economic Characters in CucumberDocument10 pagesBreeding For Fusarium Wilt Resistance and Some Economic Characters in CucumberShailendra RajanNo ratings yet
- Development of The Mass-Rearing Technique For A Predatory Mite Stratiolaelaps Scimitus (Acari - Laelapidae) Using The Double Box SystemDocument8 pagesDevelopment of The Mass-Rearing Technique For A Predatory Mite Stratiolaelaps Scimitus (Acari - Laelapidae) Using The Double Box SystemErnesto FloresNo ratings yet
- Tomato Cultivars Resistant or Susceptible To Spider Mites Differ in Their Biosynthesis and Metabolic Profile of The Monoterpenoid PathwayDocument19 pagesTomato Cultivars Resistant or Susceptible To Spider Mites Differ in Their Biosynthesis and Metabolic Profile of The Monoterpenoid PathwayWellfritz ChristiantoNo ratings yet
- Digbijaya Swain, Et AlDocument12 pagesDigbijaya Swain, Et AlDigbijaya SwainNo ratings yet
- Rayburn Genome SizeDocument6 pagesRayburn Genome SizeSebastian DavisNo ratings yet
- Molecular identification of bean genotypes for partial resistance to Sclerotinia sclerotiorum based on SCAR markersDocument7 pagesMolecular identification of bean genotypes for partial resistance to Sclerotinia sclerotiorum based on SCAR markersGleice LimaNo ratings yet
- Pfge Genotyping of Metallo Beta Lactamase Producing Pseudomonas AeruginosaDocument5 pagesPfge Genotyping of Metallo Beta Lactamase Producing Pseudomonas AeruginosaIJAR JOURNALNo ratings yet
- Antifungal Activity Against Plant Pathogenic Fungi of Chaetoviridins Isolated From Chaetomium GlobosumDocument5 pagesAntifungal Activity Against Plant Pathogenic Fungi of Chaetoviridins Isolated From Chaetomium GlobosummaczweiNo ratings yet
- Admin Vol35 2iDocument6 pagesAdmin Vol35 2iAja TourayNo ratings yet
- Population Genetic Variation Analysis of Bitter GoDocument11 pagesPopulation Genetic Variation Analysis of Bitter GoCarolinaNo ratings yet
- Fritz2006 PDFDocument7 pagesFritz2006 PDFMarichuAlvaradoNo ratings yet
- VitaletalDocument6 pagesVitaletalElgie JimenezNo ratings yet
- Gene Pyramiding QTL To RD6Document8 pagesGene Pyramiding QTL To RD6Fortis's GuitarmanNo ratings yet
- tmp43EC TMPDocument7 pagestmp43EC TMPFrontiersNo ratings yet
- Clone SpecificDocument10 pagesClone SpecificKaren LizethNo ratings yet
- Paula Et Al 2021Document13 pagesPaula Et Al 2021Debora Pires PaulaNo ratings yet
- Abo-State Et Al., 2015Document7 pagesAbo-State Et Al., 2015Jordana KalineNo ratings yet
- Ujmr 2020 - 4 - 24 - 30 PDFDocument7 pagesUjmr 2020 - 4 - 24 - 30 PDFUMYU Journal of Microbiology Research (UJMR)No ratings yet
- Alimudin GMR22Document10 pagesAlimudin GMR22Rosyida Ismi (Rosyi)No ratings yet
- 13 108 Ijat2009 - 88FDocument9 pages13 108 Ijat2009 - 88FNilasari Martha DewiNo ratings yet
- Pyramiding SCN Resistance AllelesDocument10 pagesPyramiding SCN Resistance AllelesArlam SilvaNo ratings yet
- Sequence Related Amplified Polymorphism (SRAP) Marker, Potato, Bacterial Wilt.Document9 pagesSequence Related Amplified Polymorphism (SRAP) Marker, Potato, Bacterial Wilt.Linh NguyenNo ratings yet
- Genetic Mapping of RYMV Resistance and Relationship to Rice MorphologyDocument14 pagesGenetic Mapping of RYMV Resistance and Relationship to Rice MorphologyCleaver BrightNo ratings yet
- Theobroma Cacao PhytophthoraDocument10 pagesTheobroma Cacao PhytophthoraOpenaccess Research paperNo ratings yet
- Emma - Pdis 06 22 1350 SCDocument3 pagesEmma - Pdis 06 22 1350 SCOlajide Emmanuel OlorunfemiNo ratings yet
- Control Biológico Erwinia 1Document7 pagesControl Biológico Erwinia 1Gabriela SanchezNo ratings yet
- Multilocus Differentiation of The Related Dermatophytes Microsporum Canis, Microsporum Ferrugineum and Microsporum AudouiniiDocument7 pagesMultilocus Differentiation of The Related Dermatophytes Microsporum Canis, Microsporum Ferrugineum and Microsporum AudouiniiFina Rahmatul UmmahNo ratings yet
- Wahyudi Et AlDocument7 pagesWahyudi Et AlShilajit BaruaNo ratings yet
- ECOLOGICAL-GENETICDocument8 pagesECOLOGICAL-GENETICMickaël Houessou SononNo ratings yet
- Evaluation of Rhizosphere Bacterial Antagonists For Their Potential To BioprotectDocument7 pagesEvaluation of Rhizosphere Bacterial Antagonists For Their Potential To BioprotectHvzem MohammedNo ratings yet
- Exploring The PKS and NRPS Gene From Egyptian SoilDocument11 pagesExploring The PKS and NRPS Gene From Egyptian SoilfajarakiraNo ratings yet
- Crop Protectants 2015Document9 pagesCrop Protectants 2015joyeeta8No ratings yet
- Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and GeneticsFrom EverandPlant Resistance to Herbivores and Pathogens: Ecology, Evolution, and GeneticsNo ratings yet
- A Comprehensive Analysis On Spread andDocument14 pagesA Comprehensive Analysis On Spread andcata castroNo ratings yet
- Evidence For Host-Specificity of Culturable Fungal Root Endophytes From The Carnivorous Plant Pinguicula Vulgaris (Common Butterwort)Document3 pagesEvidence For Host-Specificity of Culturable Fungal Root Endophytes From The Carnivorous Plant Pinguicula Vulgaris (Common Butterwort)Raymundo Lopez NNo ratings yet
- Agentes Causales de Faciacion y ProliferacionDocument10 pagesAgentes Causales de Faciacion y ProliferacionHugo Zagal MaldonadoNo ratings yet
- Mapping Powdery Mildew Resistance Gene pmYBL on Chromosome 7BDocument30 pagesMapping Powdery Mildew Resistance Gene pmYBL on Chromosome 7BperunicaNo ratings yet
- Lysin - Fmicb 13 856473Document11 pagesLysin - Fmicb 13 856473Sudeepta SinghNo ratings yet
- Density-Dependent Prophylaxis in The Mealworm Beetle Tenebrio Molitor L. (Coleoptera: Tenebrionidae) : Cuticular Melanization Is An Indicator of Investment in ImmunityDocument6 pagesDensity-Dependent Prophylaxis in The Mealworm Beetle Tenebrio Molitor L. (Coleoptera: Tenebrionidae) : Cuticular Melanization Is An Indicator of Investment in ImmunityajudNo ratings yet
- Nicotine's Defensive Function in Nature: Open Access, Freely Available Online BiologyDocument7 pagesNicotine's Defensive Function in Nature: Open Access, Freely Available Online BiologyAnang Yanuar RamadhanNo ratings yet
- Impact of Diazotrophic Bacteria On Germination and Growth of Tomato, With Bio-Control Effect, Isolated From Algerian SoilDocument7 pagesImpact of Diazotrophic Bacteria On Germination and Growth of Tomato, With Bio-Control Effect, Isolated From Algerian SoilsaidaNo ratings yet
- MarkerDocument8 pagesMarkershyama_deviNo ratings yet
- Ahn 2005 AllelopathyDocument7 pagesAhn 2005 AllelopathyTran Dang KhanhNo ratings yet
- Systemic Resistance Induced in Arabidopsis Thaliana by Trichoderma Asperellum SKT-1, A Microbial Pesticide of Seed-Borne Diseases of RiceDocument22 pagesSystemic Resistance Induced in Arabidopsis Thaliana by Trichoderma Asperellum SKT-1, A Microbial Pesticide of Seed-Borne Diseases of RicePrineteejayNo ratings yet
- 2 46 1652348513 6ijbrjun202206Document6 pages2 46 1652348513 6ijbrjun202206TJPRC PublicationsNo ratings yet
- Fiber-Optic Communications System: The Basic BlocksDocument9 pagesFiber-Optic Communications System: The Basic BlocksKilari SrigowriNo ratings yet
- Design and Analysis of Three-Stage Amplifier For Driving pF-to-nF Capacitive Load Based On Local Compensation TechniquesDocument18 pagesDesign and Analysis of Three-Stage Amplifier For Driving pF-to-nF Capacitive Load Based On Local Compensation TechniquesKilari SrigowriNo ratings yet
- Design and Analysis of Three-Stage Amplifier For Driving pF-to-nF Capacitive Load Based On Local Compensation TechniquesDocument18 pagesDesign and Analysis of Three-Stage Amplifier For Driving pF-to-nF Capacitive Load Based On Local Compensation TechniquesKilari SrigowriNo ratings yet
- Scattering Loss and Bending LossDocument2 pagesScattering Loss and Bending LossKilari SrigowriNo ratings yet
- Optical Sources and DetectorsDocument30 pagesOptical Sources and DetectorsKilari SrigowriNo ratings yet
- Color Thresholding Method For Image Segmentation o PDFDocument8 pagesColor Thresholding Method For Image Segmentation o PDFKilari SrigowriNo ratings yet
- Color Thresholding Method For Image Segmentation o PDFDocument8 pagesColor Thresholding Method For Image Segmentation o PDFKilari SrigowriNo ratings yet
- EC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & SciencesDocument19 pagesEC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & SciencesKilari SrigowriNo ratings yet
- EC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & ScienceDocument24 pagesEC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & ScienceKilari SrigowriNo ratings yet
- EC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & ScienceDocument24 pagesEC 415 - VLSI Design: Anil Neerukonda Institute of Tech. & ScienceKilari SrigowriNo ratings yet
- 5.a Personal Diet Consultant For Healthy MealDocument5 pages5.a Personal Diet Consultant For Healthy MealKishore SahaNo ratings yet
- Staining TechniquesDocument31 pagesStaining TechniquesKhadija JaraNo ratings yet
- 10 Tips To Support ChildrenDocument20 pages10 Tips To Support ChildrenRhe jane AbucejoNo ratings yet
- C11 Strategy DevelopmentDocument30 pagesC11 Strategy DevelopmentPARTI KEADILAN RAKYAT NIBONG TEBALNo ratings yet
- Continuous Sampling PlanDocument2 pagesContinuous Sampling PlanAnonymous wA6NGuyklDNo ratings yet
- Banu Maaruf of The LevantDocument6 pagesBanu Maaruf of The LevantMotiwala AbbasNo ratings yet
- Simulia Abaqus Standard DatasheetDocument3 pagesSimulia Abaqus Standard Datasheetuser923019231831No ratings yet
- The Interview: P F T IDocument14 pagesThe Interview: P F T IkkkkccccNo ratings yet
- Final - WPS PQR 86Document4 pagesFinal - WPS PQR 86Parag WadekarNo ratings yet
- English Extra Conversation Club International Women's DayDocument2 pagesEnglish Extra Conversation Club International Women's Dayevagloria11No ratings yet
- Delhi (The Capital of India) : Ebook by Ssac InstituteDocument27 pagesDelhi (The Capital of India) : Ebook by Ssac InstituteAnanjay ChawlaNo ratings yet
- How To Pass in University Arrear Exams?Document26 pagesHow To Pass in University Arrear Exams?arrearexam100% (6)
- SeparatorDocument2 pagesSeparatormmk111No ratings yet
- Oracle® E-Business Suite: Integrated SOA Gateway Implementation Guide Release 12.2Document202 pagesOracle® E-Business Suite: Integrated SOA Gateway Implementation Guide Release 12.2yadavdevenderNo ratings yet
- Decembermagazine 2009Document20 pagesDecembermagazine 2009maria_diyah4312No ratings yet
- CP Mother's Day Lesson PlanDocument2 pagesCP Mother's Day Lesson PlanAma MiriNo ratings yet
- POPULARITY OF CREDIT CARDS ISSUED BY DIFFERENT BANKSDocument25 pagesPOPULARITY OF CREDIT CARDS ISSUED BY DIFFERENT BANKSNaveed Karim Baksh75% (8)
- PAPERBOARD QUALITYDocument8 pagesPAPERBOARD QUALITYaurelia carinaNo ratings yet
- People v. Lagarto, 326 SCRA 693 (2000)Document43 pagesPeople v. Lagarto, 326 SCRA 693 (2000)GioNo ratings yet
- United Airlines Case Study: Using Marketing to Address External ChallengesDocument4 pagesUnited Airlines Case Study: Using Marketing to Address External ChallengesSakshiGuptaNo ratings yet
- Pebc CompilationDocument14 pagesPebc CompilationAarti AroraNo ratings yet
- 3.1 C 4.5 Algorithm-19Document10 pages3.1 C 4.5 Algorithm-19nayan jainNo ratings yet
- Virtio-Fs - A Shared File System For Virtual MachinesDocument21 pagesVirtio-Fs - A Shared File System For Virtual MachinesLeseldelaterreNo ratings yet
- Analysis and Design of Multi Storey Building by Using STAAD ProDocument5 pagesAnalysis and Design of Multi Storey Building by Using STAAD Prolikith rockNo ratings yet
- Building and Enhancing New Literacies Across The CurriculumDocument119 pagesBuilding and Enhancing New Literacies Across The CurriculumLowela Kasandra100% (3)
- Oral Medication PharmacologyDocument4 pagesOral Medication PharmacologyElaisa Mae Delos SantosNo ratings yet
- Communications201101 DLDocument132 pagesCommunications201101 DLAle SandovalNo ratings yet
- Basic LCI To High LCIDocument3 pagesBasic LCI To High LCIIonut VladNo ratings yet
- PrintDocument18 pagesPrintHookandcrookNo ratings yet
- Operating Manual: Please Read These Operating Instructions Before Using Your FreedomchairDocument24 pagesOperating Manual: Please Read These Operating Instructions Before Using Your FreedomchairNETHYA SHARMANo ratings yet