You might also like
- Skeletal Muscle Tissue in Movement and Health: Positives and NegativesDocument6 pagesSkeletal Muscle Tissue in Movement and Health: Positives and NegativesSeri RahmadaniNo ratings yet
- Muscle Biophysics - From Molecules To Cells - D. RassieerDocument374 pagesMuscle Biophysics - From Molecules To Cells - D. RassieerGigiEnergieNo ratings yet
- Physiology and BiochemistryFrom EverandPhysiology and BiochemistryGeoffrey BourneNo ratings yet
- Review Do Muscles Function As Adaptable Locomotor SpringsDocument6 pagesReview Do Muscles Function As Adaptable Locomotor SpringsItai IzhakNo ratings yet
- Muscle Fascia and StrengthDocument5 pagesMuscle Fascia and StrengthPedro CorrêaNo ratings yet
- J MusculoskPainDocument4 pagesJ MusculoskPainBruno OliveiraNo ratings yet
- Biomechanism of StretchingDocument11 pagesBiomechanism of Stretchingsara mohamedNo ratings yet
- Muscle ContractionDocument8 pagesMuscle Contractionwx8vnnnzjhNo ratings yet
- Muscular Contraction HuxleyDocument20 pagesMuscular Contraction HuxleyJersonFabianMaldonadoMorenoNo ratings yet
- 1 s2.0 S0021929023001525 MainDocument12 pages1 s2.0 S0021929023001525 MainJose PerezNo ratings yet
- Anatomy and Human Movement: Structure and FunctionFrom EverandAnatomy and Human Movement: Structure and FunctionRating: 3.5 out of 5 stars3.5/5 (2)
- Lindstedt 2001Document6 pagesLindstedt 2001Bruno Marques StreyNo ratings yet
- The Journal of Physiology, Vol. 243, No. 1 Frontispiece: HuxleyDocument45 pagesThe Journal of Physiology, Vol. 243, No. 1 Frontispiece: HuxleyTakis StefanopoulosNo ratings yet
- The Biomechanics of Stretching: Duane KnudsonDocument10 pagesThe Biomechanics of Stretching: Duane KnudsonAnuragNo ratings yet
- The Biomechanics of Stretching: Duane KnudsonDocument10 pagesThe Biomechanics of Stretching: Duane KnudsonSanjanaNo ratings yet
- Ijans - Understanding Static Muscular Contractions and Bodily MovementsDocument6 pagesIjans - Understanding Static Muscular Contractions and Bodily Movementsiaset123No ratings yet
- Core StabilityDocument4 pagesCore StabilityWhisperer BowenNo ratings yet
- Functional and Clinical Significance of Skeletal Muscle ArchitectureDocument20 pagesFunctional and Clinical Significance of Skeletal Muscle Architectureapi-3702726No ratings yet
- Skeletal Muscle Mechanics: Questions, Problems and Possible SolutionsDocument17 pagesSkeletal Muscle Mechanics: Questions, Problems and Possible Solutionskombe mukobeNo ratings yet
- Human KinesiologyDocument26 pagesHuman KinesiologyTrinidad Farías PiñeiroNo ratings yet
- APTITU FISICA - Metter - Mortalidad y FuerzaDocument9 pagesAPTITU FISICA - Metter - Mortalidad y FuerzaDVD FUTBOLNo ratings yet
- BHATTACHARYAet - Al OccupationalErgonomics PrinciplesandApplicationschapter104PATTYsTOXHANDBOOK2010 PDFDocument39 pagesBHATTACHARYAet - Al OccupationalErgonomics PrinciplesandApplicationschapter104PATTYsTOXHANDBOOK2010 PDFJorge Luis Raygada AzpilcuetaNo ratings yet
- Book Reviews: Muscle DamageDocument3 pagesBook Reviews: Muscle DamageManisha SahNo ratings yet
- Craniofacial Biomechanics An Overview of Recent Multibody Modeling StudiesDocument10 pagesCraniofacial Biomechanics An Overview of Recent Multibody Modeling StudiesDenise MathreNo ratings yet
- Trappe 2004Document13 pagesTrappe 2004cristianNo ratings yet
- Muscle Structure, Fiber Types, and Physiology: January 2015Document32 pagesMuscle Structure, Fiber Types, and Physiology: January 2015JUAN CORTES ESPINOSANo ratings yet
- 05 - Journal of Anatomy - 2013 - GerlingDocument11 pages05 - Journal of Anatomy - 2013 - Gerlingalan.consultoriafitnessNo ratings yet
- Solomomow 2006 Ligamento y Control MotorDocument19 pagesSolomomow 2006 Ligamento y Control MotorBeto RebolledoNo ratings yet
- Infant Oli No 2012Document7 pagesInfant Oli No 2012HeculesNo ratings yet
- RG7601B45 - Homework 2 - BTY632 - MANISH 3040060045Document6 pagesRG7601B45 - Homework 2 - BTY632 - MANISH 3040060045Jaskirat SinghNo ratings yet
- Journal Of: Architectural Properties of The First Dorsal Interosseous MuscleDocument7 pagesJournal Of: Architectural Properties of The First Dorsal Interosseous MuscleHeculesNo ratings yet
- MusclereviewfinalDocument14 pagesMusclereviewfinalAvelino De Leon B. JrNo ratings yet
- Skeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Document14 pagesSkeletal Muscle: A Brief Review of Structure and Function: Calcified Tissue International October 2014Anonymous K9tmd7T0PzNo ratings yet
- Mechanical Properties of Respiratory MusclesDocument69 pagesMechanical Properties of Respiratory Musclescard breatheNo ratings yet
- The Action of Two-Joint MusclesDocument16 pagesThe Action of Two-Joint MusclesDolly SuriyaamaritNo ratings yet
- Narici Et Al. 2003 - EnvelhecimentoDocument7 pagesNarici Et Al. 2003 - EnvelhecimentoFábio LanferdiniNo ratings yet
- Fascial or Muscle StretchingDocument11 pagesFascial or Muscle StretchingClinica paliativosNo ratings yet
- Muscles Within Muscles Coordination of 1Document17 pagesMuscles Within Muscles Coordination of 1luiz henrique matiasNo ratings yet
- Research Paper On MusclesDocument5 pagesResearch Paper On Musclesgz91rnat100% (1)
- Biotensegrity-The Mechanics of Fascia: December 2012Document11 pagesBiotensegrity-The Mechanics of Fascia: December 2012cacho2000No ratings yet
- Stretch-Induced Enhancement of Mechanical PowerDocument8 pagesStretch-Induced Enhancement of Mechanical PowerCharisNo ratings yet
- Anatomical Note Spurt and Shunt Muscles: MadeDocument3 pagesAnatomical Note Spurt and Shunt Muscles: MadeDonato FormicolaNo ratings yet
- 16 FullDocument11 pages16 FullEdwin VillavicencioNo ratings yet
- Muscular Adaptations and Insulin-Like Growth FactoDocument13 pagesMuscular Adaptations and Insulin-Like Growth Factoleal thiagoNo ratings yet
- 30-Second Anatomy: The 50 most important structures and systems in the human body each explained in under half a minuteFrom Everand30-Second Anatomy: The 50 most important structures and systems in the human body each explained in under half a minuteRating: 2 out of 5 stars2/5 (1)
- The History Dependence of Force Production in Mammalian Skeletal Muscle Following Stretch-Shortening and Shortening-Stretch CyclesDocument12 pagesThe History Dependence of Force Production in Mammalian Skeletal Muscle Following Stretch-Shortening and Shortening-Stretch Cyclesapi-3702726No ratings yet
- Ates F Et Al 2019Document10 pagesAtes F Et Al 2019blaiNo ratings yet
- Experimental Physiology - Historical Perspective: Celebrating 100 Years of Publishing Discovery in PhysiologyDocument4 pagesExperimental Physiology - Historical Perspective: Celebrating 100 Years of Publishing Discovery in PhysiologyJason MeehanNo ratings yet
- Fnint 11 00002Document15 pagesFnint 11 00002labsoneducationNo ratings yet
- A Review of Fascia and Its Force Transmission CapacityDocument11 pagesA Review of Fascia and Its Force Transmission CapacityShayne ShotNo ratings yet
- DOIS Seireg 1973Document16 pagesDOIS Seireg 1973GERARDO TORRES RUIZNo ratings yet
- Factors Contributing To Lower Metabolic Demand of Eccentric Compared With Concentric CyclingDocument10 pagesFactors Contributing To Lower Metabolic Demand of Eccentric Compared With Concentric CyclingFrancisco Javier Luza RamosNo ratings yet
- Biblio - Control Motor Oyt 2015Document6 pagesBiblio - Control Motor Oyt 2015Alan L'DoppelgangerNo ratings yet
- Review Article: Force Transmission Between Synergistic Skeletal Muscles Through Connective Tissue LinkagesDocument9 pagesReview Article: Force Transmission Between Synergistic Skeletal Muscles Through Connective Tissue Linkagesrcastello20No ratings yet
- Jackson (2016)Document16 pagesJackson (2016)Bruno TeixeiraNo ratings yet
- Skeletal Muscle Fatigue PDFDocument47 pagesSkeletal Muscle Fatigue PDFSun UlfahNo ratings yet
- LAPORAN LENGKAP Jaringan OtotDocument26 pagesLAPORAN LENGKAP Jaringan Ototjeon niaNo ratings yet
- The Collected Papers of Irvin M. Korr Vol. IDocument257 pagesThe Collected Papers of Irvin M. Korr Vol. IGaël Dans100% (9)
- A Plausible Explanation of The Double-Slit Experiment in Quantum PhysicsDocument3 pagesA Plausible Explanation of The Double-Slit Experiment in Quantum PhysicsCraig MawsonNo ratings yet
- AC PPTDocument28 pagesAC PPTPrajwal BirwadkarNo ratings yet
- Unit 1Document57 pagesUnit 121ELB370MOHAMMAD AREEB HASAN KHANNo ratings yet
- CHM170L Exp6 Heat of CombustionDocument5 pagesCHM170L Exp6 Heat of CombustionKaiser SaltoNo ratings yet
- Logistic RegressionDocument10 pagesLogistic RegressionNikhil GandhiNo ratings yet
- SAP SD Interview QuestionsDocument130 pagesSAP SD Interview QuestionsPraveen Kumar100% (1)
- BS 1370 1979Document13 pagesBS 1370 1979thushtikaNo ratings yet
- ORM. Lucene. Elasticsearch. Integrated. - Hibernate SearchDocument6 pagesORM. Lucene. Elasticsearch. Integrated. - Hibernate SearchChandu ChandrakanthNo ratings yet
- World's Leading Supplier of Infrared (IR) Receivers: OptoelectronicsDocument4 pagesWorld's Leading Supplier of Infrared (IR) Receivers: OptoelectronicsHenry ChanNo ratings yet
- Ucsf ChimeraDocument10 pagesUcsf ChimeraAhmed MashalyNo ratings yet
- Green Synthesis of Copper Nanoparticles and Its CharacterizationDocument5 pagesGreen Synthesis of Copper Nanoparticles and Its CharacterizationNicolas MolinaNo ratings yet
- Electrical UnitsDocument2 pagesElectrical Unitsevtoma100% (1)
- Assignment AEP 2014Document5 pagesAssignment AEP 2014Sagar MohanNo ratings yet
- INGEPAC LINE PROTECTION RELAY-overviewDocument2 pagesINGEPAC LINE PROTECTION RELAY-overviewmilan9bNo ratings yet
- OsseointegrationDocument99 pagesOsseointegrationNitin H.c.100% (2)
- VSM Blue)Document4 pagesVSM Blue)api-27411749No ratings yet
- Unification of Euler and Werner Deconvolution in Three Dimensions Via The Generalized Hilbert TransformDocument6 pagesUnification of Euler and Werner Deconvolution in Three Dimensions Via The Generalized Hilbert TransformMithunNo ratings yet
- Design of A Floating Roof Crude Oil Storage Tank of 100,000 BPD Capacity and Prototype FabricationDocument13 pagesDesign of A Floating Roof Crude Oil Storage Tank of 100,000 BPD Capacity and Prototype FabricationMudit VermaNo ratings yet
- Curl and DivDocument3 pagesCurl and DivgjdapromiseNo ratings yet
- Che Vol1Document139 pagesChe Vol1abiramanNo ratings yet
- Simulink SimulationDocument15 pagesSimulink Simulationpoopoopho100% (1)
- 1 CathDocument12 pages1 CathHashem AlsmadiNo ratings yet
- Contaminants in Landfill SoilsDocument8 pagesContaminants in Landfill SoilsChoi KhoironNo ratings yet
- Innovative Building Material: Shrihari K. BhuskadeDocument15 pagesInnovative Building Material: Shrihari K. Bhuskademla.santoshNo ratings yet
- BalancesDocument8 pagesBalancescchauling6841No ratings yet
- Turbine Control SystemDocument8 pagesTurbine Control SystemZakariya50% (2)
- Adaptive Cruise Control (Acc)Document7 pagesAdaptive Cruise Control (Acc)bnc1No ratings yet
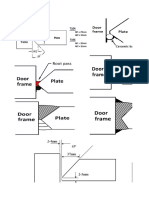
- Door SheetDocument9 pagesDoor SheetAnilkumarNo ratings yet
- Manual Montaj Cobra 600iDocument130 pagesManual Montaj Cobra 600iDdanaa DanaNo ratings yet
- VB Exercises LoopsDocument6 pagesVB Exercises LoopsS Ahmad1311993No ratings yet