You might also like
- Stem Cells and Cell Signalling in Skeletal MyogenesisFrom EverandStem Cells and Cell Signalling in Skeletal MyogenesisNo ratings yet
- Tspiralis RevDocument6 pagesTspiralis Revbumble blick crumplestickNo ratings yet
- Cell-Cell Signaling in Vertebrate DevelopmentFrom EverandCell-Cell Signaling in Vertebrate DevelopmentE.J. RobertsonRating: 5 out of 5 stars5/5 (1)
- Neural Stem Cells: Developmental Mechanisms and Disease ModelingDocument6 pagesNeural Stem Cells: Developmental Mechanisms and Disease ModelingDi RaNo ratings yet
- Forum: Neural Stem Cells and Neurogenesis in The AdultDocument3 pagesForum: Neural Stem Cells and Neurogenesis in The AdultGustavo López ToledoNo ratings yet
- Mielinizacion 2Document12 pagesMielinizacion 2sabadboy 01No ratings yet
- Currie, 2016 PDFDocument18 pagesCurrie, 2016 PDF22gaefNo ratings yet
- HHS Public Access: Origins of Glial Cell Populations in The Insect Nervous SystemDocument18 pagesHHS Public Access: Origins of Glial Cell Populations in The Insect Nervous SystemHelena QuintNo ratings yet
- Nones 2010 Flavonoids and Astrocytes Crosstalking Implications For Brain Development and PathologyDocument12 pagesNones 2010 Flavonoids and Astrocytes Crosstalking Implications For Brain Development and PathologyrosybioNo ratings yet
- 2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusDocument8 pages2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusNityananda PortelladaNo ratings yet
- FACS-based Isolation of Neural and Glioma Stem Cell Populations From Fresh Human Tissues Utilizing EGF LigandDocument23 pagesFACS-based Isolation of Neural and Glioma Stem Cell Populations From Fresh Human Tissues Utilizing EGF LigandPatricia GomesNo ratings yet
- Microglia Roles in Synaptic Plasticity and Myelination in Homeostatic Conditions and Neurodevelopmental DisordersDocument18 pagesMicroglia Roles in Synaptic Plasticity and Myelination in Homeostatic Conditions and Neurodevelopmental Disordersbia_corrêaNo ratings yet
- TMP EAB5Document6 pagesTMP EAB5FrontiersNo ratings yet
- M. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationDocument9 pagesM. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationHutsDMNo ratings yet
- 2017-@-SMAD7 Deficiency Stimulates Müller Progenitor Cell Proliferation During The Development of The Mammalian RetinaDocument12 pages2017-@-SMAD7 Deficiency Stimulates Müller Progenitor Cell Proliferation During The Development of The Mammalian RetinaReza NaghibiNo ratings yet
- Characteristics of Whale Mulle Glia in Primary and Immortalized Cultures - Pereiro 2022Document15 pagesCharacteristics of Whale Mulle Glia in Primary and Immortalized Cultures - Pereiro 2022Rebeca ChávezNo ratings yet
- Perspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Document6 pagesPerspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Fahmi IsmailNo ratings yet
- Oligodendrocyte, Astrocyte, and Microglia Crosstalk in Myelin Development, Damage, and RepairDocument16 pagesOligodendrocyte, Astrocyte, and Microglia Crosstalk in Myelin Development, Damage, and Repairkuldeep sainiNo ratings yet
- Emb Oj 2009327 ADocument14 pagesEmb Oj 2009327 Athemoonwalker2014No ratings yet
- Role of Extracellular Vesicles in Glia-Neuron Intercellular Communication - MP13Document11 pagesRole of Extracellular Vesicles in Glia-Neuron Intercellular Communication - MP13ASTRID GIANELLA QUIROZ NAVARRONo ratings yet
- Nihms 144952Document9 pagesNihms 144952angelos96papNo ratings yet
- Neurorepair and Regeneration of The Brain A DecadeDocument16 pagesNeurorepair and Regeneration of The Brain A DecadeRade PetrovicNo ratings yet
- Cell Division and Inheritance: Chapter OutlineDocument21 pagesCell Division and Inheritance: Chapter OutlineTannu BhambhuNo ratings yet
- Cell Division and Inheritance PatternDocument22 pagesCell Division and Inheritance PatternSheraz ChaudharyNo ratings yet
- Directly Converted Astrocytes Retain The Ageing Features of The Donor Fibroblasts and Elucidate The Astrocytic Contribution To Human CNS Health and Disease - Gatto - Aging Cell - Wiley Online LibraryDocument22 pagesDirectly Converted Astrocytes Retain The Ageing Features of The Donor Fibroblasts and Elucidate The Astrocytic Contribution To Human CNS Health and Disease - Gatto - Aging Cell - Wiley Online LibraryGustavo Sá MottaNo ratings yet
- 2articulo - (2023) Genetic Control of Meiosis Surveillance Mechanisms in MammalsDocument22 pages2articulo - (2023) Genetic Control of Meiosis Surveillance Mechanisms in Mammalsmauricio yucumaNo ratings yet
- Tmp827a TMPDocument23 pagesTmp827a TMPFrontiersNo ratings yet
- მიკროგლიაDocument13 pagesმიკროგლიაFiqria KhomasuridzeNo ratings yet
- Basic Concepts of EpigeneticsDocument10 pagesBasic Concepts of EpigeneticsRaju MehtaNo ratings yet
- Neural Stem Cells: Balancing Self-Renewal With DifferentiationDocument13 pagesNeural Stem Cells: Balancing Self-Renewal With DifferentiationValeria FigueroaNo ratings yet
- Review: Microglia Biology: One Century of Evolving ConceptsDocument20 pagesReview: Microglia Biology: One Century of Evolving ConceptsPiali ChatterjeeNo ratings yet
- MJA Dev Biol 2018 444, S110-S143 PDFDocument34 pagesMJA Dev Biol 2018 444, S110-S143 PDFmjaybarNo ratings yet
- Microglia-Derived Microvesicles Affect Microglia Phenotype in GliomaDocument14 pagesMicroglia-Derived Microvesicles Affect Microglia Phenotype in GliomaPaige MunroeNo ratings yet
- Transforming Growth Factor-A1 Is A Negative Modulator of Adult NeurogenesisDocument13 pagesTransforming Growth Factor-A1 Is A Negative Modulator of Adult Neurogenesisdr. RiyanNo ratings yet
- Jean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsDocument12 pagesJean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsHutsDMNo ratings yet
- García 2003 - Role of Muller Glia in Neuroprotection and Regeneration in The RetinaDocument14 pagesGarcía 2003 - Role of Muller Glia in Neuroprotection and Regeneration in The RetinaRebeca VictoriaNo ratings yet
- Fncel 11 00024Document17 pagesFncel 11 00024Eka LalaNo ratings yet
- Stem Cells: Dr. Valerie Bracchi-RicardDocument49 pagesStem Cells: Dr. Valerie Bracchi-RicardE12H12No ratings yet
- Neurogeneisis in RatsDocument7 pagesNeurogeneisis in RatsjsaccuzzoNo ratings yet
- Genes Synap Atten MSDocument6 pagesGenes Synap Atten MSjohn fossellaNo ratings yet
- Extracellular Regulators of Axonal Growth in The Adult Central Nervous SystemDocument18 pagesExtracellular Regulators of Axonal Growth in The Adult Central Nervous SystemLuis CatalánNo ratings yet
- FullDocument8 pagesFullrocambolescas perthNo ratings yet
- A Neuroscientist S Guide To Transgenic Mice An - 2020 - Neuroscience - BiobehaviDocument17 pagesA Neuroscientist S Guide To Transgenic Mice An - 2020 - Neuroscience - BiobehaviNabil Sangga BuanaNo ratings yet
- 1642 FullDocument18 pages1642 FullFahrunnisa NurdinNo ratings yet
- Local Cues Establish and Maintain Region Specific Phenotypes of Ba - 2017 - NeurDocument23 pagesLocal Cues Establish and Maintain Region Specific Phenotypes of Ba - 2017 - NeurNikita MiroshnichenkoNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- 361 13patilDocument5 pages361 13patilTanveer SultanNo ratings yet
- Essay - Nanou - Discuss New Cancer Treatment Approach - Biol - Basics - Psychol - 2021 - Fall PDFDocument9 pagesEssay - Nanou - Discuss New Cancer Treatment Approach - Biol - Basics - Psychol - 2021 - Fall PDFMaria NaNo ratings yet
- Computational Identification of Specific Genes For Glioblastoma Stem-Like Cells IdentityDocument10 pagesComputational Identification of Specific Genes For Glioblastoma Stem-Like Cells IdentityBG BBNo ratings yet
- ARDA Embryo 3rd HWDocument3 pagesARDA Embryo 3rd HWSamantha ArdaNo ratings yet
- Different Effects of Anti-Sonic Hedgehog AntibodieDocument10 pagesDifferent Effects of Anti-Sonic Hedgehog AntibodieVERONICA BLANCONo ratings yet
- Flochlay Et Al 2023Document26 pagesFlochlay Et Al 2023aaki2627No ratings yet
- EpigeneticaDocument9 pagesEpigeneticaChristian Moises CasasNo ratings yet
- Interneurons in The Developing Human Neocortex: Nada Zecevic, Frances Hu, Igor JakovcevskiDocument16 pagesInterneurons in The Developing Human Neocortex: Nada Zecevic, Frances Hu, Igor JakovcevskiFrontiersNo ratings yet
- Soares-Vieira2021 Article MicrogliaAtTheCentreOfBrainResDocument16 pagesSoares-Vieira2021 Article MicrogliaAtTheCentreOfBrainResAdriana NataliaNo ratings yet
- Generic Master ChefsDocument11 pagesGeneric Master ChefsAquib AmirNo ratings yet
- Brain Cancer Stem CellsDocument53 pagesBrain Cancer Stem CellsYinebeb MezgebuNo ratings yet
- Joa 235 432Document20 pagesJoa 235 432ElCanaldeSaadNo ratings yet
- 2020 - The Fate and Function of Oligodendrocyte Progenitor Cells After Traumatic Spinal Cord InjuryDocument19 pages2020 - The Fate and Function of Oligodendrocyte Progenitor Cells After Traumatic Spinal Cord InjuryMariana NannettiNo ratings yet
- Bianconi 2013Document10 pagesBianconi 2013Maria Júlia ToderoNo ratings yet
- Sorensen2017 PDFDocument37 pagesSorensen2017 PDFPaige MunroeNo ratings yet
- Cells: The Potential of Targeting Brain Pathology With Ascl1/Mash1Document7 pagesCells: The Potential of Targeting Brain Pathology With Ascl1/Mash1Paige MunroeNo ratings yet
- Solga2015 PDFDocument13 pagesSolga2015 PDFPaige MunroeNo ratings yet
- In Search For Reliable Markers of Glioma-Induced Polarization of MicrogliaDocument13 pagesIn Search For Reliable Markers of Glioma-Induced Polarization of MicrogliaPaige MunroeNo ratings yet
- Aberrant Expression of Interleukin-1 B and Inflammasome Activation in Human Malignant GliomasDocument14 pagesAberrant Expression of Interleukin-1 B and Inflammasome Activation in Human Malignant GliomasPaige MunroeNo ratings yet
- Microglia Immunophenotyping in Gliomas: Abstract. Microglia, Once Assimilated To Peripheral MacroDocument9 pagesMicroglia Immunophenotyping in Gliomas: Abstract. Microglia, Once Assimilated To Peripheral MacroPaige MunroeNo ratings yet
- HHS Public AccessDocument22 pagesHHS Public AccessPaige MunroeNo ratings yet
- Faria2006 PDFDocument11 pagesFaria2006 PDFPaige MunroeNo ratings yet
- Frank 2016Document21 pagesFrank 2016Paige MunroeNo ratings yet
- The oncometabolite 2-hydroxyglutarate inhibits microglial activation via the AMPK/mTOR/NF-κB pathwayDocument11 pagesThe oncometabolite 2-hydroxyglutarate inhibits microglial activation via the AMPK/mTOR/NF-κB pathwayPaige MunroeNo ratings yet
- Microglia-Derived Microvesicles Affect Microglia Phenotype in GliomaDocument14 pagesMicroglia-Derived Microvesicles Affect Microglia Phenotype in GliomaPaige MunroeNo ratings yet
- Erik Jung, Julieta Alfonso, Hannah Monyer, Wolfgang Wick, Frank WinklerDocument41 pagesErik Jung, Julieta Alfonso, Hannah Monyer, Wolfgang Wick, Frank WinklerPaige MunroeNo ratings yet
- Guo 2019Document13 pagesGuo 2019Paige MunroeNo ratings yet
- Neurochemistry International: SciencedirectDocument12 pagesNeurochemistry International: SciencedirectPaige MunroeNo ratings yet
- TLR2 Promotes Glioma Immune Evasion by Downregulating MHC Class II Molecules in MicrogliaDocument15 pagesTLR2 Promotes Glioma Immune Evasion by Downregulating MHC Class II Molecules in MicrogliaPaige MunroeNo ratings yet
- Cadherin-Dependent Adhesion of Human U373MG Glioblastoma Cells Promotes Neurite Outgrowth and Increases Migratory CapacityDocument7 pagesCadherin-Dependent Adhesion of Human U373MG Glioblastoma Cells Promotes Neurite Outgrowth and Increases Migratory CapacityPaige MunroeNo ratings yet
- Frank 2015Document27 pagesFrank 2015Paige MunroeNo ratings yet
- Tumour-Processed Osteopontin and Lactadherin Drive The Protumorigenic Reprogramming of Microglia and Glioma ProgressionDocument12 pagesTumour-Processed Osteopontin and Lactadherin Drive The Protumorigenic Reprogramming of Microglia and Glioma ProgressionPaige MunroeNo ratings yet
- Brandenburg2015 PDFDocument14 pagesBrandenburg2015 PDFPaige MunroeNo ratings yet
- Susan G. Cady2 and Masanori Sono: Archives of Biochemistry AND BiophysicsDocument8 pagesSusan G. Cady2 and Masanori Sono: Archives of Biochemistry AND BiophysicsPaige MunroeNo ratings yet
- Glioblastoma-Associated Microglia Reprogramming Is Mediated by Functional Transfer of Extracellular Mir-21Document23 pagesGlioblastoma-Associated Microglia Reprogramming Is Mediated by Functional Transfer of Extracellular Mir-21Paige MunroeNo ratings yet
- Intravital 2-Photon Imaging Reveals Distinct Morphology and Infiltrative Properties of Glioblastoma-Associated MacrophagesDocument6 pagesIntravital 2-Photon Imaging Reveals Distinct Morphology and Infiltrative Properties of Glioblastoma-Associated MacrophagesPaige MunroeNo ratings yet
- MSC Exam Spectroscopy 2018 FINALDocument2 pagesMSC Exam Spectroscopy 2018 FINALPaige MunroeNo ratings yet
- Cellular Forms and Functions of Brain Microglia: ReviewDocument6 pagesCellular Forms and Functions of Brain Microglia: ReviewPaige MunroeNo ratings yet
- CCL 107 PDFDocument3 pagesCCL 107 PDFPaige MunroeNo ratings yet
- Toxic RelationshipDocument1 pageToxic RelationshipwidyasNo ratings yet
- Cross Rate and Merchant RateDocument26 pagesCross Rate and Merchant RateDivya NadarajanNo ratings yet
- Silage Sampling and AnalysisDocument5 pagesSilage Sampling and AnalysisG_ASantosNo ratings yet
- Region Iii - Central Luzon Padapada National High SchoolDocument2 pagesRegion Iii - Central Luzon Padapada National High SchoolRotipNo ratings yet
- Bee Keeping-KVK MorenaDocument6 pagesBee Keeping-KVK MorenaAsh1Scribd100% (1)
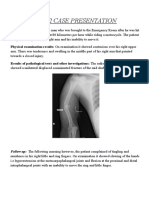
- PBL 2 Case PresentationDocument12 pagesPBL 2 Case PresentationRamish IrfanNo ratings yet
- Biology Project Cronary Heart Diseas (CHD)Document7 pagesBiology Project Cronary Heart Diseas (CHD)احمد المغربي50% (2)
- Vocab PDFDocument29 pagesVocab PDFShahab SaqibNo ratings yet
- Decision Making in Perioperative Medicine Clinical Pearls 2021Document351 pagesDecision Making in Perioperative Medicine Clinical Pearls 2021Dal RdzNo ratings yet
- Content of An Investigational New Drug Application (IND)Document13 pagesContent of An Investigational New Drug Application (IND)Prathamesh MaliNo ratings yet
- Case Study LenovoDocument10 pagesCase Study LenovoGOHAR GHAFFARNo ratings yet
- Marketing ProjectDocument82 pagesMarketing ProjectSumit GuptaNo ratings yet
- 03 Secondary School Student's Academic Performance Self Esteem and School Environment An Empirical Assessment From NigeriaDocument10 pages03 Secondary School Student's Academic Performance Self Esteem and School Environment An Empirical Assessment From NigeriaKienstel GigantoNo ratings yet
- English BeginnersDocument34 pagesEnglish BeginnersCristina ZamfirNo ratings yet
- What Does She/He Look Like?: Height Build AGEDocument18 pagesWhat Does She/He Look Like?: Height Build AGEHenrich Garcia LimaNo ratings yet
- Sithpat006ccc019 A - 2021.1Document34 pagesSithpat006ccc019 A - 2021.1Mark Andrew Clarete100% (2)
- Containers HandbookDocument26 pagesContainers Handbookrishi vohraNo ratings yet
- POB Ch08Document28 pagesPOB Ch08Anjum MalikNo ratings yet
- URICA TestDocument3 pagesURICA TestCristy PagalanNo ratings yet
- Meng Mammogram JulDocument4 pagesMeng Mammogram JulLivia MillNo ratings yet
- Margarita ForesDocument20 pagesMargarita ForesKlarisse YoungNo ratings yet
- Indirect Current Control of LCL Based Shunt Active Power FilterDocument10 pagesIndirect Current Control of LCL Based Shunt Active Power FilterArsham5033No ratings yet
- HOPE 3-Module 8-Week 5-6Document9 pagesHOPE 3-Module 8-Week 5-6Freya SalorNo ratings yet
- 7 LevelDocument5 pages7 LevelACHREF RIHANINo ratings yet
- The Preparation of Culture MediaDocument7 pagesThe Preparation of Culture MediaNakyanzi AngellaNo ratings yet
- PYMS Is A Reliable Malnutrition Screening ToolsDocument8 pagesPYMS Is A Reliable Malnutrition Screening ToolsRika LedyNo ratings yet
- WCC PSV PDFDocument40 pagesWCC PSV PDFAlejandro RamelaNo ratings yet
- Intraocular Pressure and Aqueous Humor DynamicsDocument36 pagesIntraocular Pressure and Aqueous Humor DynamicsIntan EkarulitaNo ratings yet
- Pieces 1 Cod Amount Product Detail 36 Cotton Pant RemarksDocument4 pagesPieces 1 Cod Amount Product Detail 36 Cotton Pant RemarksFaizan AhmadNo ratings yet
- Occlusal Appliance TherapyDocument14 pagesOcclusal Appliance TherapyNam BuiNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4.5 out of 5 stars4.5/5 (6)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (597)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (111)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Remnants of Ancient Life: The New Science of Old FossilsFrom EverandRemnants of Ancient Life: The New Science of Old FossilsRating: 3 out of 5 stars3/5 (3)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignFrom EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignRating: 4 out of 5 stars4/5 (19)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (116)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Civilized To Death: The Price of ProgressFrom EverandCivilized To Death: The Price of ProgressRating: 4.5 out of 5 stars4.5/5 (215)