You might also like
- Clinical Nutrition ESPEN: Hiroshige Itakura, Michie Kobayashi, Seiji NakamuraDocument7 pagesClinical Nutrition ESPEN: Hiroshige Itakura, Michie Kobayashi, Seiji NakamuraPáblo EugênioNo ratings yet
- ... Bao Et Al2015Document8 pages... Bao Et Al2015Gabriel GursenNo ratings yet
- 1 s2.0 S0271531714000724 MainDocument9 pages1 s2.0 S0271531714000724 MainNurma SabilalaNo ratings yet
- Am J Clin Nutr 2012 Theytaz 1008 16Document9 pagesAm J Clin Nutr 2012 Theytaz 1008 16evaNo ratings yet
- Dietary Fats, Carbohydrates and Vascular Disease: Sri Lankan PerspectivesDocument4 pagesDietary Fats, Carbohydrates and Vascular Disease: Sri Lankan PerspectivesViro Hernandez VilladiegoNo ratings yet
- Boss Ola 2009Document8 pagesBoss Ola 2009nettoxsNo ratings yet
- Basu 2009Document8 pagesBasu 2009abida.raufkNo ratings yet
- Ziziphus Spina Christi20200317 4568 1vzdzos With Cover Page v2Document15 pagesZiziphus Spina Christi20200317 4568 1vzdzos With Cover Page v2Rosnidar SumardiNo ratings yet
- Artigo InglesDocument8 pagesArtigo InglesViviani FlorNo ratings yet
- Jurnal TomatDocument10 pagesJurnal TomatSEPTIA SRI EKA PUTRINo ratings yet
- LycopeneDocument7 pagesLycopeneafridiNo ratings yet
- Effects of Grapefruit and Seville Orange Juices On The Pharmacokinetic Properties of Colchicine in Healthy Subjects - ScienceDirectDocument28 pagesEffects of Grapefruit and Seville Orange Juices On The Pharmacokinetic Properties of Colchicine in Healthy Subjects - ScienceDirectRoy BisentiNo ratings yet
- Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular RiskDocument10 pagesIntestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular RiskLeonardoNo ratings yet
- Nihms 1550215Document27 pagesNihms 1550215muh. almusyafirNo ratings yet
- Macronutrient Intake Induces Oxidative and Inflammatory Stress Potential Relevance To Atherosclerosis and Insulin ResistanceDocument9 pagesMacronutrient Intake Induces Oxidative and Inflammatory Stress Potential Relevance To Atherosclerosis and Insulin ResistanceJohn RonquilloNo ratings yet
- Emerging Applications of Metabolomics ToDocument20 pagesEmerging Applications of Metabolomics ToMariela GonzalezNo ratings yet
- Lycopene and Heart HealthDocument8 pagesLycopene and Heart HealthDr. HeleNo ratings yet
- ChoiDocument19 pagesChoiLuciana RafaelNo ratings yet
- Artificially Cultivated Ophiocordyceps Sinensis AlleviatesDocument17 pagesArtificially Cultivated Ophiocordyceps Sinensis AlleviatesHoang IceNo ratings yet
- Targeting The NLRP3 Inflammasome To Reduce Diet-Induced Metabolic Abnormalities in MiceDocument39 pagesTargeting The NLRP3 Inflammasome To Reduce Diet-Induced Metabolic Abnormalities in Miceanirbanmanna88320No ratings yet
- Pca 3086Document14 pagesPca 3086Piyush BhallaNo ratings yet
- The Effect of Drinking Water PH On The Human Gut Microbiota and Glucose Regulation: Results of A Randomized Controlled Cross-Over InterventionDocument12 pagesThe Effect of Drinking Water PH On The Human Gut Microbiota and Glucose Regulation: Results of A Randomized Controlled Cross-Over InterventionFlor Da PrimaveraNo ratings yet
- Biomolecules 11 00937Document9 pagesBiomolecules 11 00937Nasrin SharifiNo ratings yet
- 2010 Article 9216 PDFDocument8 pages2010 Article 9216 PDFphoechoexNo ratings yet
- 2016 Physical Activity and Liver DiseasesDocument15 pages2016 Physical Activity and Liver Diseasesorellanaroco.10No ratings yet
- Am J Clin Nutr 2014 Lecoultre 268 75Document8 pagesAm J Clin Nutr 2014 Lecoultre 268 75Kang Opik TaufikNo ratings yet
- Diet and TherapyDocument3 pagesDiet and TherapyKatty AlevonNo ratings yet
- BiomedicinesDocument12 pagesBiomedicinessamuelNo ratings yet
- Estadella Et Al, 2013Document14 pagesEstadella Et Al, 2013Daniela OliveiraNo ratings yet
- Adv Nutr-2011-Hyson-408-20Document13 pagesAdv Nutr-2011-Hyson-408-20Lia CoandrasNo ratings yet
- A Moderate-Fat Diet With One Avocado Per Day Increases Plasma Antioxidants and Decreases The Oxidation of Small, Dense LDL in Adults With Overweight and Obesity RCT 2019Document9 pagesA Moderate-Fat Diet With One Avocado Per Day Increases Plasma Antioxidants and Decreases The Oxidation of Small, Dense LDL in Adults With Overweight and Obesity RCT 2019Kelly DickinsonNo ratings yet
- Article: Correspondence: Lrobinso@uoguelph - Ca Tel.: +1-519-824-4120 (Ext. 52297)Document16 pagesArticle: Correspondence: Lrobinso@uoguelph - Ca Tel.: +1-519-824-4120 (Ext. 52297)Alma AcevedoNo ratings yet
- Low - and High - Doses of EtOH Insulin Sensitivity 2007Document10 pagesLow - and High - Doses of EtOH Insulin Sensitivity 2007Gál Bence SzabóNo ratings yet
- Jci37385 PDFDocument13 pagesJci37385 PDFNur Ahmad HabibiNo ratings yet
- Glucose As A Major Antioxidant - When, What For and Why It FailsDocument20 pagesGlucose As A Major Antioxidant - When, What For and Why It FailsDaniel GilbertoNo ratings yet
- Ijo 201493 ADocument9 pagesIjo 201493 AKurtisNo ratings yet
- Probióticos para Tratamiento de Hígado Graso No AlcohólicoDocument8 pagesProbióticos para Tratamiento de Hígado Graso No AlcohólicoNorlan Ariel Caldera NavarreteNo ratings yet
- Srep11276+ +Omega+3+e+MicrobiotaDocument17 pagesSrep11276+ +Omega+3+e+MicrobiotaVanessa BarrosNo ratings yet
- Flaxseed OilDocument36 pagesFlaxseed OilKamal UddinNo ratings yet
- Zidani 2016Document30 pagesZidani 2016Pembawa KebahagiaanNo ratings yet
- Anti-Inflammatory E Polymeric Nanoparticles: Relevance of Particle Internalization in Endothelial CellsDocument17 pagesAnti-Inflammatory E Polymeric Nanoparticles: Relevance of Particle Internalization in Endothelial CellsRika DestianaNo ratings yet
- Ajtr0002 0316Document16 pagesAjtr0002 0316bagas umam alwiNo ratings yet
- Hyperglycemia in Critical Illness: A ReviewDocument11 pagesHyperglycemia in Critical Illness: A ReviewSreejith ViswanathanNo ratings yet
- Soal Latihan PartografDocument9 pagesSoal Latihan PartografADHITYA YUDHA MAULANANo ratings yet
- Flavanoids and EDDocument8 pagesFlavanoids and EDstarmittNo ratings yet
- Patel 2020Document31 pagesPatel 2020larissa brandaliseNo ratings yet
- Hep4 1 726Document10 pagesHep4 1 726Medicina InternaNo ratings yet
- Jurnal Bahasa InggrisDocument6 pagesJurnal Bahasa InggrisDepi Murdiansyah100% (2)
- Tjelle Grape Juice Blood PressureDocument10 pagesTjelle Grape Juice Blood PressureLJ NissenNo ratings yet
- Review of Literature: Chapter - 2Document46 pagesReview of Literature: Chapter - 2InaGargNo ratings yet
- Effect of Supplementation With Polyphenol Extract of Thymus Lipid Profile in High-Fat Diet - Fed HamstersDocument11 pagesEffect of Supplementation With Polyphenol Extract of Thymus Lipid Profile in High-Fat Diet - Fed Hamstersbiodoc biodocNo ratings yet
- Diet Rich in High Glucoraphanin Broccoli Reduces Plasma LDL Cholesterol: Evidence From Randomised Controlled TrialsDocument9 pagesDiet Rich in High Glucoraphanin Broccoli Reduces Plasma LDL Cholesterol: Evidence From Randomised Controlled TrialsYuni SahoeNo ratings yet
- De La Rosa 2015Document12 pagesDe La Rosa 2015yasmin matterNo ratings yet
- KersenDocument13 pagesKersenJumasni AdnanNo ratings yet
- Moon 2011Document9 pagesMoon 2011Nia Karomatul AuliaNo ratings yet
- Bioavailability and Antioxidant Effects of Orange Juice Components in HumansDocument9 pagesBioavailability and Antioxidant Effects of Orange Juice Components in HumansjohnheverthNo ratings yet
- Fish Oil in Rheumatic Diseases.Document8 pagesFish Oil in Rheumatic Diseases.José Isaac Lamas GarcíaNo ratings yet
- Immunomodulation by Lipid Emulsions in Pulmonary Inflammation A Randomized Controlled TrialDocument13 pagesImmunomodulation by Lipid Emulsions in Pulmonary Inflammation A Randomized Controlled TrialMartyn PereiraNo ratings yet
- High Density Lipoproteins, Dyslipidemia, and Coronary Heart DiseaseFrom EverandHigh Density Lipoproteins, Dyslipidemia, and Coronary Heart DiseaseNo ratings yet
- Evaluasi Program Pemberian Makanan Tambahan Pemulihan (PMT-P) Terhadap Status Gizi Buruk Balita Di Puskesmas Oepoi Kota KupangDocument8 pagesEvaluasi Program Pemberian Makanan Tambahan Pemulihan (PMT-P) Terhadap Status Gizi Buruk Balita Di Puskesmas Oepoi Kota KupangLeni PurbasariNo ratings yet
- Pengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia PrasekolahDocument9 pagesPengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia Prasekolahsentaro2No ratings yet
- Efektifitas Program Pemberian Makanan Tambahan-Pemulihan Pada Ibu Hamil Kurang Energi Kronik Di Kota PalembangDocument10 pagesEfektifitas Program Pemberian Makanan Tambahan-Pemulihan Pada Ibu Hamil Kurang Energi Kronik Di Kota PalembangUmbu JhenNo ratings yet
- Evaluasi Program Pemberian Makanan Tambahan Pemulihan (PMT-P) Terhadap Status Gizi Buruk Balita Di Puskesmas Oepoi Kota KupangDocument8 pagesEvaluasi Program Pemberian Makanan Tambahan Pemulihan (PMT-P) Terhadap Status Gizi Buruk Balita Di Puskesmas Oepoi Kota KupangLeni PurbasariNo ratings yet
- Am J Clin Nutr-2004-Aljada-682-90-1Document9 pagesAm J Clin Nutr-2004-Aljada-682-90-1Leni PurbasariNo ratings yet
- Peningkatan Higiene Sanitasi Sebagai Upaya Menjaga Kualitas Makanan Dan Kepuasan Pelanggan Di Rumah Makan Dhamar PalembangDocument9 pagesPeningkatan Higiene Sanitasi Sebagai Upaya Menjaga Kualitas Makanan Dan Kepuasan Pelanggan Di Rumah Makan Dhamar PalembangNova Dwi LestariNo ratings yet
- Pola AsuhDocument5 pagesPola AsuhOnil Wakil Tuhan100% (1)
- Pola AsuhDocument5 pagesPola AsuhOnil Wakil Tuhan100% (1)
- Pola AsuhDocument5 pagesPola AsuhOnil Wakil Tuhan100% (1)
- Pengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia PrasekolahDocument9 pagesPengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia Prasekolahsentaro2No ratings yet
- Pengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia PrasekolahDocument9 pagesPengaruh Penyuluhan Gizi-Kesehatan Dan Faktor Lainnya Terhadap Pertumbuhan Anak Usia Prasekolahsentaro2No ratings yet
- Nutrition and Chronic ConditionsDocument228 pagesNutrition and Chronic ConditionsLeni PurbasariNo ratings yet
- 4 5920233068844549631Document442 pages4 5920233068844549631Indahtul MufidahNo ratings yet
- MAGGINI 2007 Selected Vitamins, Trace Element and Immune FunctionDocument7 pagesMAGGINI 2007 Selected Vitamins, Trace Element and Immune FunctionLeni PurbasariNo ratings yet
- Cage HomesDocument8 pagesCage HomesstanhopekrisNo ratings yet
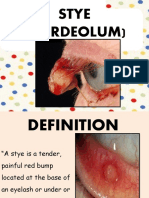
- StyeDocument21 pagesStyenur syafiqah kamaruzamanNo ratings yet
- Maxime Gorky and KingstonDocument433 pagesMaxime Gorky and KingstonBasil FletcherNo ratings yet
- Information On The Use of Domperidone To Increase Milk Production in Lactating WomenDocument3 pagesInformation On The Use of Domperidone To Increase Milk Production in Lactating WomenKhairul HananNo ratings yet
- Actisorb Silver 220: Product ProfileDocument12 pagesActisorb Silver 220: Product ProfileBeaulah HunidzariraNo ratings yet
- Drug StudyDocument11 pagesDrug StudyKimberly Subade MandilagNo ratings yet
- Tendonitis Guide - Causes, Symptoms and Treatment OptionsDocument6 pagesTendonitis Guide - Causes, Symptoms and Treatment OptionsSylvia GraceNo ratings yet
- Nowadays Young People Spend Too Much of Their Free Time in Shopping MallsDocument3 pagesNowadays Young People Spend Too Much of Their Free Time in Shopping MallsYacine BenHmeidaNo ratings yet
- National Geographic USA - January 2016Document148 pagesNational Geographic USA - January 2016stamenkovskib100% (4)
- Human Body Systems: Study GuideDocument11 pagesHuman Body Systems: Study Guideapi-242114183No ratings yet
- Blood Can Be Very BadDocument37 pagesBlood Can Be Very BadPhil SingerNo ratings yet
- PrioritizationDocument4 pagesPrioritizationJhennie EyescandieNo ratings yet
- A Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaDocument8 pagesA Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaEditor IJTSRDNo ratings yet
- Systema Let Every BreathDocument82 pagesSystema Let Every BreathB Agel100% (1)
- Environmental Studies-FIRST UNIT-vkmDocument59 pagesEnvironmental Studies-FIRST UNIT-vkmRandomNo ratings yet
- Agarwal - Manual of Neuro OphthalmogyDocument272 pagesAgarwal - Manual of Neuro Ophthalmogythycoon100% (4)
- Pathophysiology of Ischemic Stroke FinalDocument3 pagesPathophysiology of Ischemic Stroke FinalAcohCChao67% (3)
- COPD CurrentDocument9 pagesCOPD Currentmartha kurniaNo ratings yet
- HEX PG SyllabusDocument124 pagesHEX PG SyllabusJegadeesan MuniandiNo ratings yet
- Subjective: Diarrhea Related To Watery Short Term: IndependentDocument4 pagesSubjective: Diarrhea Related To Watery Short Term: IndependentEmma Lyn SantosNo ratings yet
- 8mkebook PDFDocument300 pages8mkebook PDFVanja Vujovic100% (2)
- NUR091 - Session 8Document6 pagesNUR091 - Session 8aquinoemjay27No ratings yet
- Menstrual Blood Derived Stem Cells and Their Scope in Regenerative Medicine A Review ArticleDocument6 pagesMenstrual Blood Derived Stem Cells and Their Scope in Regenerative Medicine A Review ArticleInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Management of Imature ApexDocument21 pagesManagement of Imature ApexBogdanNo ratings yet
- Zoology MCQs Practice Test 19Document4 pagesZoology MCQs Practice Test 19Rehan HaiderNo ratings yet
- Cast and SplintsDocument58 pagesCast and SplintsSulabh Shrestha100% (2)
- Sri Lanka - Averting A National Nutrition Anomaly 2Document16 pagesSri Lanka - Averting A National Nutrition Anomaly 2Rushan LakdimuthuNo ratings yet
- MBF90324234Document407 pagesMBF90324234Emad Mergan100% (1)
- GMO Pro Arguments (Gen-WPS OfficeDocument4 pagesGMO Pro Arguments (Gen-WPS OfficeFranchesca RevelloNo ratings yet
- Criminal SociologyDocument53 pagesCriminal SociologyJackielyn cabayaoNo ratings yet