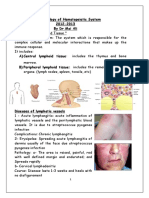
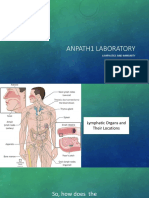
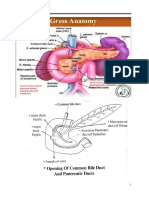
You might also like
- Department of Histology, Cytology and EmbryologyDocument8 pagesDepartment of Histology, Cytology and Embryologysubmen81No ratings yet
- Órganos Linfoides TerciariosDocument8 pagesÓrganos Linfoides Terciariosjenith andradeNo ratings yet
- 4 2Document3 pages4 2Stefia AisyahNo ratings yet
- Cardiovascular LymphaticDocument17 pagesCardiovascular LymphaticFaisal Ridho SaktiNo ratings yet
- Anatomy& EmbryologyDocument13 pagesAnatomy& EmbryologySravani PeddagangannagariNo ratings yet
- Detmar Onc2011602aDocument10 pagesDetmar Onc2011602aChetan PatilNo ratings yet
- Lymph SystemDocument7 pagesLymph SystemghaiathNo ratings yet
- Vanmeter: Gould'S Pathophysiology For The Health Professions, 5Th EditionDocument1 pageVanmeter: Gould'S Pathophysiology For The Health Professions, 5Th EditionAnne VaughanNo ratings yet
- Histology - Immune and Hemopoietic OrgansDocument23 pagesHistology - Immune and Hemopoietic OrgansKacper DaraszkiewiczNo ratings yet
- Chapter 22 Summary Chapter 22 SummaryDocument13 pagesChapter 22 Summary Chapter 22 SummarytinecastilloNo ratings yet
- Pathology of Hematopoietic System 2012-2013 by DR - Mai AliDocument17 pagesPathology of Hematopoietic System 2012-2013 by DR - Mai AliMuhammad Hilman Abu BakarNo ratings yet
- Anatomy - Cardiovascular System - Part IIIDocument12 pagesAnatomy - Cardiovascular System - Part IIIVăn ĐứcNo ratings yet
- Histology 10 ImmunosystemDocument79 pagesHistology 10 ImmunosystemAbdul RahmanNo ratings yet
- Lymphatics and BloodDocument9 pagesLymphatics and BloodIchokwu VivianNo ratings yet
- (DD13-14) Anatomic Sciences PDFDocument861 pages(DD13-14) Anatomic Sciences PDFminiarNo ratings yet
- Case VirchowsDocument14 pagesCase VirchowsShanti IntansariNo ratings yet
- Basic Immunology PDFDocument6 pagesBasic Immunology PDFAnastasia ENo ratings yet
- Anatomy of Lymphatics SystemDocument9 pagesAnatomy of Lymphatics Systemtg856qr4mwNo ratings yet
- The Lymphatic SystemDocument7 pagesThe Lymphatic SystemPreeti ChouhanNo ratings yet
- Laporan Kasus VI Bahasa InggrisDocument10 pagesLaporan Kasus VI Bahasa InggrisWahyu SholekhuddinNo ratings yet
- Fisiologia LinfedemaDocument8 pagesFisiologia LinfedemaSAKAI69No ratings yet
- LymphaticDocument3 pagesLymphaticTania Louise Pioquinto AbuanNo ratings yet
- Lymphocyte TraffickingDocument8 pagesLymphocyte TraffickingAbhishek KumarNo ratings yet
- Patho - 4th Asessment - Lymphoproliferative Disorders I - 31 Jan 2007Document24 pagesPatho - 4th Asessment - Lymphoproliferative Disorders I - 31 Jan 2007api-3703352100% (1)
- Human Anatomy,: First Edition Mckinley & O'LoughlinDocument40 pagesHuman Anatomy,: First Edition Mckinley & O'LoughlinAgung PurmanaNo ratings yet
- Lymphatics BD BloodDocument10 pagesLymphatics BD BloodIchokwu VivianNo ratings yet
- Organs of The Immune SystemDocument19 pagesOrgans of The Immune SystemprabuNo ratings yet
- The Lymphatic System and Immunity by Navarro and Orjalo BS Psy 3 1Document27 pagesThe Lymphatic System and Immunity by Navarro and Orjalo BS Psy 3 1markNo ratings yet
- Lymphatic Drainage of HeadDocument142 pagesLymphatic Drainage of HeadVijayendra kamath100% (2)
- (DD13-14) Anatomic SciencesDocument861 pages(DD13-14) Anatomic SciencesIndu C. R100% (2)
- Lymphatic Drainage of Head & NeckDocument96 pagesLymphatic Drainage of Head & NeckRoopa PatilNo ratings yet
- Lymphatic SystemDocument41 pagesLymphatic SystemKemoy Francis100% (3)
- The Lymphatics in Kidney Health and Disease: Michael D. Donnan, Yael Kenig-Kozlovsky and Susan E. QuagginDocument21 pagesThe Lymphatics in Kidney Health and Disease: Michael D. Donnan, Yael Kenig-Kozlovsky and Susan E. QuagginsilviaNo ratings yet
- Immune Cells and OrgansDocument8 pagesImmune Cells and OrgansCharlz AriolaNo ratings yet
- Notes 20Document3 pagesNotes 20Paris ShaShaNo ratings yet
- Functions:: Four Functions of SpleenDocument5 pagesFunctions:: Four Functions of SpleenRashid DayaoNo ratings yet
- Magnetic Resonance LymphangiographyDocument14 pagesMagnetic Resonance LymphangiographyHector Hernandez-SoriaNo ratings yet
- For Each of The Following Multiple Choice Questions, Select The BEST AnswerDocument6 pagesFor Each of The Following Multiple Choice Questions, Select The BEST AnswerAbdul Ghaffar AbdullahNo ratings yet
- A&P Exam 2 ReviewDocument7 pagesA&P Exam 2 ReviewKathleen SaucedoNo ratings yet
- Normal Structure of Lymph Nodes & Their DistributionDocument33 pagesNormal Structure of Lymph Nodes & Their DistributionMadhan Govindaswamy GNo ratings yet
- Exercise 10 The Lymphatic System: ZOO 115 Animal Histology Laboratory Exercises 1Document4 pagesExercise 10 The Lymphatic System: ZOO 115 Animal Histology Laboratory Exercises 1Christi Lorraine LayogNo ratings yet
- 08 - Lymphatic SystemDocument19 pages08 - Lymphatic SystemDanielSenpai69No ratings yet
- Anatomy of The Lymphatic and Immune SystemsDocument5 pagesAnatomy of The Lymphatic and Immune SystemsHebsiba PonnayyanNo ratings yet
- 08 - Lymphatic SystemDocument19 pages08 - Lymphatic SystemKonstantinos KosmidisNo ratings yet
- Lymphatic System: Prof. Shekhar TeliDocument19 pagesLymphatic System: Prof. Shekhar TeliShekhar TeliNo ratings yet
- Laboratory Exercise - LymphaticsDocument5 pagesLaboratory Exercise - LymphaticsJohn Henry G. Gabriel IVNo ratings yet
- Oliver 2004Document11 pagesOliver 2004SDNKSJNDJKNo ratings yet
- Anpath1 Laboratory: Lymphatics and ImmunityDocument15 pagesAnpath1 Laboratory: Lymphatics and ImmunityYancy BagsaoNo ratings yet
- Biology Research WorkDocument4 pagesBiology Research WorkAsif HussainNo ratings yet
- The Anatomy of The Immune System (Lymphatic SystemDocument4 pagesThe Anatomy of The Immune System (Lymphatic SystemMohammed R.HusseinNo ratings yet
- Lymphatic System: By-Victor BawiahDocument42 pagesLymphatic System: By-Victor BawiahTheophilus BaidooNo ratings yet
- Lymphotropic Therapy For Diseases of The Maxillofacial Region Review of The LiteratureDocument6 pagesLymphotropic Therapy For Diseases of The Maxillofacial Region Review of The LiteratureEditor IJTSRDNo ratings yet
- H.Histology Lec1Document10 pagesH.Histology Lec1Marci MunirNo ratings yet
- Reticuloendothelial SystemDocument26 pagesReticuloendothelial SystemNashreen QtqtNo ratings yet
- Histo DR ZahraDocument56 pagesHisto DR Zahraali.mohammed.official.7No ratings yet
- DgpqmeghjkDocument2 pagesDgpqmeghjkManikandanNo ratings yet
- Lymphatic SystemDocument13 pagesLymphatic SystemCharli ParachinniNo ratings yet
- Transplant ImmunologyFrom EverandTransplant ImmunologyXian C. LiNo ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesFrom EverandCellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesCharles G. CochraneNo ratings yet
- A Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisFrom EverandA Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisNo ratings yet
- Internet of Things AssignmentDocument1 pageInternet of Things AssignmentKhadija RanaNo ratings yet
- Clean and Green PakistanDocument4 pagesClean and Green PakistanKhadija RanaNo ratings yet
- Mam MehwishDocument13 pagesMam MehwishKhadija RanaNo ratings yet
- Khadija Rana Bsee-19071 Communication Skills 2 Assignment 1: Compare and Contrast of The SpeechesDocument2 pagesKhadija Rana Bsee-19071 Communication Skills 2 Assignment 1: Compare and Contrast of The SpeechesKhadija RanaNo ratings yet
- Lymphangitis LymphadenitisDocument5 pagesLymphangitis LymphadenitisGabriela Cocieru MotelicaNo ratings yet
- MCI FMGE Previous Year Solved Question Paper 2005 PDFDocument32 pagesMCI FMGE Previous Year Solved Question Paper 2005 PDFRuby Bhola100% (1)
- White Blood Cells and Its Disorders: Key PointsDocument24 pagesWhite Blood Cells and Its Disorders: Key PointsskNo ratings yet
- Pathophysiology Notes 5Document5 pagesPathophysiology Notes 5Scotty BanksNo ratings yet
- (03des) Slide LymphedemaDocument52 pages(03des) Slide LymphedemaAbigaleNo ratings yet
- Cupping Therapy ExplainedDocument16 pagesCupping Therapy ExplainedAbdul Rahman Khan100% (3)
- Carcinoma of PancreasDocument15 pagesCarcinoma of Pancreasعمار عارفNo ratings yet
- BodyBallancer Brochure PDFDocument2 pagesBodyBallancer Brochure PDFRoss GaleNo ratings yet
- Hernia and Acute Abdomen MCQDocument44 pagesHernia and Acute Abdomen MCQShriyansh Chahar50% (4)
- Clinicals ENTDocument70 pagesClinicals ENTMarrow Pg42No ratings yet
- Hematology Handouts PDFDocument11 pagesHematology Handouts PDFJann ericka JaoNo ratings yet
- BSI Natures Self DefenceDocument12 pagesBSI Natures Self DefencesreenitthiNo ratings yet
- Anatomy and Physiology of DengueDocument4 pagesAnatomy and Physiology of Dengueciarkai_18No ratings yet
- Fun Facts About Anatomy and Physiology 2Document22 pagesFun Facts About Anatomy and Physiology 2Theodore Liwongan0% (1)
- Immunology and SerologyDocument225 pagesImmunology and SerologyialegnaNo ratings yet
- Chapter 33 Guyton and HallDocument4 pagesChapter 33 Guyton and Hallg_komolafeNo ratings yet
- First Exam Website QuestionsDocument45 pagesFirst Exam Website QuestionsSawsan AtiehNo ratings yet
- Immune Response in The Human BodyDocument30 pagesImmune Response in The Human BodyjohnosborneNo ratings yet
- Nursing Care of The Client With CancerDocument15 pagesNursing Care of The Client With CancerAlvin M Alcayno0% (1)
- Anexo-Unit1-Body SystemsDocument5 pagesAnexo-Unit1-Body SystemsPau GuauNo ratings yet
- Hodgkin Lymphoma: Dr. G Sirisha M.D Assistant Professor Department of Pathology, GMC, SrikakulamDocument41 pagesHodgkin Lymphoma: Dr. G Sirisha M.D Assistant Professor Department of Pathology, GMC, SrikakulamDevisriNo ratings yet
- LymphadenopathyDocument18 pagesLymphadenopathyIin RosalinaeNo ratings yet
- Lymphatic System and Immunity: Student Learning OutcomesDocument8 pagesLymphatic System and Immunity: Student Learning Outcomeslily1liang-1No ratings yet
- Score Report - Immunology & Serology at Free Online Quiz SchoolDocument29 pagesScore Report - Immunology & Serology at Free Online Quiz SchoolThomas GiangNo ratings yet
- 1st Term Exam Lecture NCM 106Document7 pages1st Term Exam Lecture NCM 106Marvick Galima100% (2)
- Adv Anat Pathol Volume 24, Number 6, November 2017Document25 pagesAdv Anat Pathol Volume 24, Number 6, November 2017María Fernanda EstévezNo ratings yet
- Lymph Drainage Therapy PDFDocument7 pagesLymph Drainage Therapy PDFDiego Mamani Mullisaca100% (2)
- JawapanLengkap Modul Biology PDFDocument32 pagesJawapanLengkap Modul Biology PDFT-specialist SNo ratings yet
- LimfadenopatiDocument7 pagesLimfadenopatiLestari Santika DewiNo ratings yet
- Limfangitis Nur AmaliaDocument17 pagesLimfangitis Nur AmaliaIka Lukita SariNo ratings yet