You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Strategies, Techniques, Approaches To Critical Thinking A Clinical Reasoning Workbook For Nurses, 5e by Sandra Luz Martinez de Castillo EdD RN PDFDocument287 pagesStrategies, Techniques, Approaches To Critical Thinking A Clinical Reasoning Workbook For Nurses, 5e by Sandra Luz Martinez de Castillo EdD RN PDFJ1525% (8)
- NAC Protocol PrintableDocument21 pagesNAC Protocol Printablemaadyyy123100% (1)
- Pharmacology of AntibioticDocument66 pagesPharmacology of AntibioticfgrehNo ratings yet
- Traveler's DiarrheaDocument17 pagesTraveler's DiarrheaAnonymous D29e00100% (1)
- Clinical LaboratoriesDocument24 pagesClinical Laboratoriespdamodar200788% (8)
- Atrial Septal Defect (ASD)Document35 pagesAtrial Septal Defect (ASD)Nur Arifah Astri100% (2)
- Case Pres-Banag Laum SuperfinaleDocument46 pagesCase Pres-Banag Laum SuperfinaleAyen FornollesNo ratings yet
- Lymphatic System of The Dog BaumDocument219 pagesLymphatic System of The Dog BaumFelipeeNo ratings yet
- Nama Riwayat PekerjaanDocument35 pagesNama Riwayat PekerjaanfgrehNo ratings yet
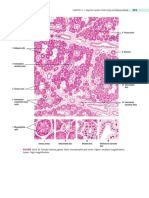
- CHAPTER 11 - Digestive System: Oral Cavity and Salivary GlandsDocument3 pagesCHAPTER 11 - Digestive System: Oral Cavity and Salivary GlandsfgrehNo ratings yet
- Artigo Caries EstevamDocument7 pagesArtigo Caries EstevamfgrehNo ratings yet
- Dosen Wali Utk Di Cek MahasiswaDocument28 pagesDosen Wali Utk Di Cek MahasiswafgrehNo ratings yet
- Jadwal Skill'S Lab (Revisi) SEMESTER GANJIL T. A. 2020/2021Document23 pagesJadwal Skill'S Lab (Revisi) SEMESTER GANJIL T. A. 2020/2021fgrehNo ratings yet
- BMR - Dhita Dalilla Akrab X Mipa 10 PDFDocument3 pagesBMR - Dhita Dalilla Akrab X Mipa 10 PDFfgrehNo ratings yet
- Gigi Molar 1 Maksila Permanen: Mesiobuccal Root Distobuccal Root Lingual RootDocument3 pagesGigi Molar 1 Maksila Permanen: Mesiobuccal Root Distobuccal Root Lingual RootfgrehNo ratings yet
- Pharmacology of Anaesthetics: DR - Siti Syarifah, MbiomedDocument28 pagesPharmacology of Anaesthetics: DR - Siti Syarifah, MbiomedfgrehNo ratings yet
- Ultrastructural Gigi U PraktikumDocument64 pagesUltrastructural Gigi U PraktikumfgrehNo ratings yet
- Elisa - PCR - Electrophoresa: Dr. Malayana R Nasution, SPPKDocument30 pagesElisa - PCR - Electrophoresa: Dr. Malayana R Nasution, SPPKfgrehNo ratings yet
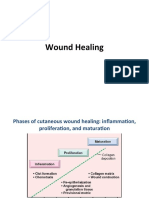
- Wound HealingDocument15 pagesWound HealingfgrehNo ratings yet
- Sistem Saraf Perifer: Dept. Fisiologi Fakultas Kedokteran USUDocument31 pagesSistem Saraf Perifer: Dept. Fisiologi Fakultas Kedokteran USUfgrehNo ratings yet
- Superteeen PDFDocument1 pageSuperteeen PDFfgrehNo ratings yet
- Sensasi Somatik: Nyeri: Dept. Fisiologi FK USUDocument22 pagesSensasi Somatik: Nyeri: Dept. Fisiologi FK USUfgrehNo ratings yet
- Praktikum 2Document6 pagesPraktikum 2fgrehNo ratings yet
- 1anesthesia For Day Care Surgery 123Document49 pages1anesthesia For Day Care Surgery 123Parvathy R NairNo ratings yet
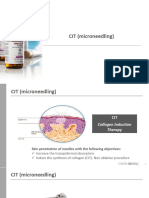
- Microneedling & Nanopore PresentationDocument58 pagesMicroneedling & Nanopore PresentationkatherineNo ratings yet
- Bacterial KeratitisDocument31 pagesBacterial KeratitisMuhammad Rehan SarfrazNo ratings yet
- Large CystDocument4 pagesLarge CystKin DacikinukNo ratings yet
- Early Childhood Caries Epidemiology, Aetiology, Risk Assessment, Societal Burden, Management, Education, and Policy: Global PerspectiveDocument11 pagesEarly Childhood Caries Epidemiology, Aetiology, Risk Assessment, Societal Burden, Management, Education, and Policy: Global PerspectiveAbdul RasheedNo ratings yet
- Anthropometrics Measurement 1 1Document7 pagesAnthropometrics Measurement 1 1Quencee CalaycayNo ratings yet
- Catalogo Linea Coronaria 2021Document8 pagesCatalogo Linea Coronaria 2021RichiNo ratings yet
- LAS HEALTH 6 Q4 Wk5 Cherlyn. RonquilloDocument4 pagesLAS HEALTH 6 Q4 Wk5 Cherlyn. RonquilloANGELINA RAMBOYONGNo ratings yet
- CTK-USA (Aria) : End User Price List-Rapid TestDocument2 pagesCTK-USA (Aria) : End User Price List-Rapid TestAbdalazeez AlsayedNo ratings yet
- Anaerobic InfectionsDocument57 pagesAnaerobic Infectionsentistde0% (1)
- Paediatric DKA Integrated Care Pathway For CportalDocument39 pagesPaediatric DKA Integrated Care Pathway For Cportalteresita vargasNo ratings yet
- BSCPT DU Syllabus-NITOR 4th BatchDocument23 pagesBSCPT DU Syllabus-NITOR 4th BatchHasan Rahman100% (2)
- Journal of Behavior Therapy and Experimental PsychiatryDocument7 pagesJournal of Behavior Therapy and Experimental PsychiatryBella ZulitaNo ratings yet
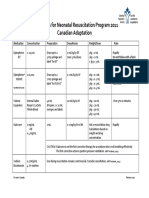
- Medications For Neonatal Resuscitation Program 2011 Canadian AdaptationDocument1 pageMedications For Neonatal Resuscitation Program 2011 Canadian AdaptationrubymayNo ratings yet
- What Causes CancerDocument2 pagesWhat Causes CancerSilla, KyshiaNo ratings yet
- P LCR Parameter Platelet Large Cell Ratio Medical Laboratories PDFDocument3 pagesP LCR Parameter Platelet Large Cell Ratio Medical Laboratories PDFluxsura8217No ratings yet
- BMJ Avian FluDocument49 pagesBMJ Avian Flualiakbar178No ratings yet
- Eye Exams For ChildrenDocument5 pagesEye Exams For ChildrenDanielz FranceNo ratings yet
- Care For Client With Oxygenation Problem: OutcomesDocument19 pagesCare For Client With Oxygenation Problem: OutcomesPaul JacksonNo ratings yet
- Aninditha. Buku Ajar Neurologi. Jakarta: Departemen Neurologi Fakultas Kedokteran Universitas Indonesia RS Cipto Mangunkusumo. 2017Document2 pagesAninditha. Buku Ajar Neurologi. Jakarta: Departemen Neurologi Fakultas Kedokteran Universitas Indonesia RS Cipto Mangunkusumo. 2017pelangiNo ratings yet
- CTG Analyzer: A Graphical User Interface For CardiotocographyDocument4 pagesCTG Analyzer: A Graphical User Interface For CardiotocographySebastian PerezNo ratings yet
- Pathophysiology Precipitating Factors: Predisposing FactorsDocument2 pagesPathophysiology Precipitating Factors: Predisposing FactorsChristine Pialan SalimbagatNo ratings yet
- Gingival Diseases in Childhood - A Review: Abst TDocument4 pagesGingival Diseases in Childhood - A Review: Abst TMagfhiro Rangga AndalasNo ratings yet