You might also like
- Comprehensive Exam ReviewDocument35 pagesComprehensive Exam ReviewIndia91% (92)
- MCU 2021 The Ventilator Book 3rd EditionDocument280 pagesMCU 2021 The Ventilator Book 3rd EditionAlvaro Estupiñan100% (2)
- Individual Health ProfileDocument2 pagesIndividual Health ProfilendpNo ratings yet
- BSM M104 Curriculum OutlineDocument3 pagesBSM M104 Curriculum OutlineDamaris Pandora Songcal0% (1)
- J Appl Physiol 2003 Heenan 2321 7Document8 pagesJ Appl Physiol 2003 Heenan 2321 7konyekNo ratings yet
- Effects of Longevinex (Modified Resveratrol) On Cardioprotection and Its Mechanisms of ActionDocument9 pagesEffects of Longevinex (Modified Resveratrol) On Cardioprotection and Its Mechanisms of ActionrhabarberrhabarberNo ratings yet
- DHEA Supplementation Improves Follicular Microenviroment in Poor Responder PatientsDocument5 pagesDHEA Supplementation Improves Follicular Microenviroment in Poor Responder Patientsomarguti795No ratings yet
- Hypobaric Hypoxia Causes Impairment of Spermatogenesis in Developing Rats at Pre-PubertyDocument7 pagesHypobaric Hypoxia Causes Impairment of Spermatogenesis in Developing Rats at Pre-PubertychristianNo ratings yet
- 1 ChestDocument5 pages1 ChestZakiyah Nur HaqqiNo ratings yet
- Effects of Prenatal Stress On Conduction Airway MorphologyDocument9 pagesEffects of Prenatal Stress On Conduction Airway MorphologyEileen Torres CerdaNo ratings yet
- Biol Sex Differ 2020 11 1 15Document10 pagesBiol Sex Differ 2020 11 1 15Fernando SousaNo ratings yet
- Urban 2005Document5 pagesUrban 2005Miyyada AichaouiNo ratings yet
- Dex 246Document12 pagesDex 246Nur WahyuniNo ratings yet
- TORCH Salmeterol Fluticasona en EpocDocument15 pagesTORCH Salmeterol Fluticasona en EpocAgustina AndradeNo ratings yet
- Pathophysiological Interpretation of Fetal Heart Rate Tracings in Clinical Practice AJOGDocument23 pagesPathophysiological Interpretation of Fetal Heart Rate Tracings in Clinical Practice AJOGMitsury Jacqueline Flores C.No ratings yet
- Lukaszuk Et Al. - 2005 - Optimization of Estradiol Supplementation During The Luteal Phase Improves The Pregnancy Rate in Women UnderDocument5 pagesLukaszuk Et Al. - 2005 - Optimization of Estradiol Supplementation During The Luteal Phase Improves The Pregnancy Rate in Women UnderKrzysztof ŁukaszukNo ratings yet
- Early and Late Analysis of Postpartum Stress in Newborn FoalsDocument8 pagesEarly and Late Analysis of Postpartum Stress in Newborn FoalsRafidah HelmiNo ratings yet
- Adrenal Androgens in Gender Affirming HTDocument2 pagesAdrenal Androgens in Gender Affirming HTJuanita LNo ratings yet
- Karatsoreos Et Al 2010Document11 pagesKaratsoreos Et Al 2010zacharymweil100% (1)
- Manning ScoreDocument10 pagesManning ScoreYurike Natalie LengkongNo ratings yet
- PIIS00150282990060Document4 pagesPIIS00150282990060lilou.magnaboscoNo ratings yet
- 2006_Publicación final BRDocument8 pages2006_Publicación final BREva Montserrat MadridNo ratings yet
- Classification and Pharmacology of ProgestinsDocument10 pagesClassification and Pharmacology of ProgestinsancaNo ratings yet
- Diet and IGF-I levels in European womenDocument9 pagesDiet and IGF-I levels in European womenYafanita IzzatiNo ratings yet
- Acos y 17b Estradiol PDFDocument22 pagesAcos y 17b Estradiol PDFAli de RoNo ratings yet
- ArticuloDocument7 pagesArticuloDulceEspinozaLNo ratings yet
- 10 1016@j Ajog 2020 06 037Document20 pages10 1016@j Ajog 2020 06 037kairalmurielNo ratings yet
- A Key Circulatory Defence Against Asphyxia in Infancy - The Heart of The Matter!Document9 pagesA Key Circulatory Defence Against Asphyxia in Infancy - The Heart of The Matter!Yasmin Salwa LestariNo ratings yet
- Fechner 2001Document7 pagesFechner 2001Daniel PmNo ratings yet
- When Is It OK to Sedate a Child Who Just AteDocument7 pagesWhen Is It OK to Sedate a Child Who Just AteNADIA VICUÑANo ratings yet
- PIIS0002937820311923Document3 pagesPIIS0002937820311923FebbyNo ratings yet
- Complementary Therapies in Clinical PracticeDocument9 pagesComplementary Therapies in Clinical PracticeKurnia NuriNo ratings yet
- tmp39EC TMPDocument3 pagestmp39EC TMPFrontiersNo ratings yet
- Neuroprotective Effects of Resveratrol on Prenatal Stress in RatsDocument12 pagesNeuroprotective Effects of Resveratrol on Prenatal Stress in RatsShobhitNo ratings yet
- Global REACH 2018: Volume Regulation in High-Altitude Andeans With and Without Chronic Mountain SicknessDocument9 pagesGlobal REACH 2018: Volume Regulation in High-Altitude Andeans With and Without Chronic Mountain SicknessFiorella LeyvaNo ratings yet
- Steroids For Airway EdemaDocument7 pagesSteroids For Airway EdemaDamal An NasherNo ratings yet
- 2005 - O'Donovan - Dose Response at On CholesterolDocument7 pages2005 - O'Donovan - Dose Response at On CholesterolFrancisco Javier Luza RamosNo ratings yet
- Extract Inhibits The Development of Hypertension in L-NAME-induced Hypertensive RatsDocument9 pagesExtract Inhibits The Development of Hypertension in L-NAME-induced Hypertensive RatsPloy PloypailinNo ratings yet
- 2008_Chamy_Biol Res -sometido-07-11-04P-PDFDocument17 pages2008_Chamy_Biol Res -sometido-07-11-04P-PDFEva Montserrat MadridNo ratings yet
- Effect of Daily Intake of Yoghurt and Bread Enriched With Biologically Active Substances On Blood Lipids and Vitamin A in Adolescents and Young AdultsDocument9 pagesEffect of Daily Intake of Yoghurt and Bread Enriched With Biologically Active Substances On Blood Lipids and Vitamin A in Adolescents and Young AdultsEsmeralda SantillanNo ratings yet
- Infertility and Eating Disorders YmobDocument1 pageInfertility and Eating Disorders Ymobps.paulayanezNo ratings yet
- Hemodynamic Gestational Adaptation in BitchesDocument6 pagesHemodynamic Gestational Adaptation in BitchesJhonatan PinheiroNo ratings yet
- Decrease in Paco2 With Prone Position Is Predictive of Improved Outcome in Acute Respiratory Distress SyndromeDocument7 pagesDecrease in Paco2 With Prone Position Is Predictive of Improved Outcome in Acute Respiratory Distress SyndromedarwigNo ratings yet
- The Use of Acupuncture With in Vitro Fertilization: Is There A Point?Document10 pagesThe Use of Acupuncture With in Vitro Fertilization: Is There A Point?lu salviaNo ratings yet
- Anesthesia For The Pregnant Cat and DogDocument5 pagesAnesthesia For The Pregnant Cat and DogDiajeng Galuh RiesantiNo ratings yet
- Betametasona y DexametasonaDocument5 pagesBetametasona y DexametasonaJmamouraNo ratings yet
- PhysicalTreatments v4n3p169 enDocument6 pagesPhysicalTreatments v4n3p169 enJulenda CintarinovaNo ratings yet
- Effects of Imposed Pursed-Lips Breathing On Respiratory Mechanics and Dyspnea at Rest and During Exercise in COPDDocument11 pagesEffects of Imposed Pursed-Lips Breathing On Respiratory Mechanics and Dyspnea at Rest and During Exercise in COPDGaoudam NatarajanNo ratings yet
- Waist Change Over 11 Years Predicts Elevated InflammationDocument6 pagesWaist Change Over 11 Years Predicts Elevated InflammationSenthamizhan PanneerNo ratings yet
- Jurnal Buaya 1Document9 pagesJurnal Buaya 1Norita RNo ratings yet
- Thorax 1981 Weiss 878 9Document3 pagesThorax 1981 Weiss 878 9Andina KluniariNo ratings yet
- Effects of Leukotrienes in The Placental VasculatuDocument2 pagesEffects of Leukotrienes in The Placental Vasculatups.paulayanezNo ratings yet
- Influence of Aerobic Exercise Training On Cardiovascular and Endocrine Inflammatory Biomarkers in Hypertensive Postmenopausal WomenDocument7 pagesInfluence of Aerobic Exercise Training On Cardiovascular and Endocrine Inflammatory Biomarkers in Hypertensive Postmenopausal WomenNemo SecretNo ratings yet
- Nihms 333002Document16 pagesNihms 333002German FloresNo ratings yet
- Dor 2Document10 pagesDor 2Alce Alce EverdienNo ratings yet
- Art:10.1007/s00360 011 0591 1Document12 pagesArt:10.1007/s00360 011 0591 1Danilo MarinNo ratings yet
- J Pediatr 2010 Mar 156 (3) 382-7Document9 pagesJ Pediatr 2010 Mar 156 (3) 382-7Hugo Velázquez GarciaNo ratings yet
- Hypertensive Disorders of Pregnancy and Long-TermDocument1 pageHypertensive Disorders of Pregnancy and Long-Termdw21541No ratings yet
- Role of VEGF in Preeclampsia PathophysiologyDocument7 pagesRole of VEGF in Preeclampsia PathophysiologyLintang SuroyaNo ratings yet
- Effect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsDocument8 pagesEffect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsBrian ColborneNo ratings yet
- Epilepsy DietDocument36 pagesEpilepsy Diettamy_001No ratings yet
- Fact #1: Wellness & Fitness-Body OptimizationDocument13 pagesFact #1: Wellness & Fitness-Body OptimizationCheeken CharliNo ratings yet
- Fetal-Placental DisordersFrom EverandFetal-Placental DisordersNicholas S. AssaliRating: 5 out of 5 stars5/5 (1)
- Hormones and the Fetus: Volume 1: Production, Concentration and Metabolism During PregnancyFrom EverandHormones and the Fetus: Volume 1: Production, Concentration and Metabolism During PregnancyNo ratings yet
- An Update On COVID-19 and Pregnancy: Djamieson@emory - EduDocument10 pagesAn Update On COVID-19 and Pregnancy: Djamieson@emory - EduRaul DoctoNo ratings yet
- 1998 Volvo s70-v70Document52 pages1998 Volvo s70-v70Raul DoctoNo ratings yet
- BMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PDocument13 pagesBMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PRaul DoctoNo ratings yet
- Satoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFDocument44 pagesSatoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFLuisNo ratings yet
- Seminar: Jan Claassen, Soojin ParkDocument17 pagesSeminar: Jan Claassen, Soojin ParkRaul DoctoNo ratings yet
- Central Venous Pressure (CVP)Document3 pagesCentral Venous Pressure (CVP)Raul DoctoNo ratings yet
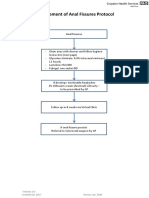
- Anal Fissures PDF 14 Sept-115042Document1 pageAnal Fissures PDF 14 Sept-115042Raul DoctoNo ratings yet
- GINA Coming SoonDocument1 pageGINA Coming SoonRaul DoctoNo ratings yet
- X3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutDocument3 pagesX3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutRaul DoctoNo ratings yet
- 1998 Volvo S70-V70Document52 pages1998 Volvo S70-V70cosminubNo ratings yet
- 2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureDocument167 pages2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureRaul DoctoNo ratings yet
- Ascoping Review of The Pathophysiology of COVID-19Document16 pagesAscoping Review of The Pathophysiology of COVID-19Raul DoctoNo ratings yet
- Rapid Diagnostic Testing For SARS-CoV-2Document9 pagesRapid Diagnostic Testing For SARS-CoV-2Wilfredo NuñezNo ratings yet
- Why Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadDocument15 pagesWhy Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadRaul Docto100% (1)
- Obstetrics & GynecologyDocument10 pagesObstetrics & GynecologyRaul DoctoNo ratings yet
- How To Install Google Play On The Amazon Fire HD 8Document7 pagesHow To Install Google Play On The Amazon Fire HD 8Raul DoctoNo ratings yet
- Robba2021 Article BasicUltrasoundHead-to-toeSkilDocument21 pagesRobba2021 Article BasicUltrasoundHead-to-toeSkilRaul DoctoNo ratings yet
- Complications of HysterectomyDocument17 pagesComplications of HysterectomyRaul DoctoNo ratings yet
- Standardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Document8 pagesStandardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Raul DoctoNo ratings yet
- AJOG Implementation of A Conservative ChecklistDocument9 pagesAJOG Implementation of A Conservative ChecklistRaul DoctoNo ratings yet
- Understanding The Role of Host Response in Influenza PneumonitisDocument3 pagesUnderstanding The Role of Host Response in Influenza PneumonitisRaul DoctoNo ratings yet
- Vlaar2021 Article TransfusionStrategiesInBleedinDocument25 pagesVlaar2021 Article TransfusionStrategiesInBleedinRaul DoctoNo ratings yet
- Case 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaDocument7 pagesCase 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaRaul DoctoNo ratings yet
- Alcohol y Metabolismo HumanoDocument21 pagesAlcohol y Metabolismo HumanoDani BelloNo ratings yet
- Surviving Sepsis Campaign International.21Document81 pagesSurviving Sepsis Campaign International.21jamesteryNo ratings yet
- Tugwell 1985Document13 pagesTugwell 1985Raul DoctoNo ratings yet
- A Framework For Standardized Management of Intrapartum PDFDocument6 pagesA Framework For Standardized Management of Intrapartum PDFRaul A UrbinaNo ratings yet
- 2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisDocument144 pages2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisSara MendesNo ratings yet
- 2014-08-13 Gynae Positioning Presentation - SlidesDocument14 pages2014-08-13 Gynae Positioning Presentation - SlidesIrina MincuNo ratings yet
- Ectopic PregnancyDocument4 pagesEctopic PregnancySyahriana AnaNo ratings yet
- Mid Im 2020 AnatomyDocument178 pagesMid Im 2020 AnatomyXzailheene Dela FuenteNo ratings yet
- Literature Review On Legalization of AbortionDocument6 pagesLiterature Review On Legalization of Abortiontkpmzasif100% (1)
- Key Facts Menstrual CycleDocument8 pagesKey Facts Menstrual CycleMari FeNo ratings yet
- NOTICE (Revised Syllabi) : (WRITTEN TEST & SYLLABI For The Posts - Advt. No. 01, 02 and 03 of 2016)Document123 pagesNOTICE (Revised Syllabi) : (WRITTEN TEST & SYLLABI For The Posts - Advt. No. 01, 02 and 03 of 2016)wani ashNo ratings yet
- Humulin R: Regular U-500 (Concentrated) Insulin Human Injection, Usp (Rdna Origin)Document7 pagesHumulin R: Regular U-500 (Concentrated) Insulin Human Injection, Usp (Rdna Origin)Rebecca BlackburnNo ratings yet
- Child Development Semester 1Document2 pagesChild Development Semester 1api-291715250No ratings yet
- An Overview of AbortionDocument60 pagesAn Overview of AbortionDivisi FER MalangNo ratings yet
- NHMS2016ReportVolumeII MaternalChildHealthFindingsv2Document276 pagesNHMS2016ReportVolumeII MaternalChildHealthFindingsv2TaufiqNo ratings yet
- APGODocument49 pagesAPGOiamdonutNo ratings yet
- Longcaseexaminationforphaseiiimedicalstudentsusmkk 100606040208 Phpapp01Document89 pagesLongcaseexaminationforphaseiiimedicalstudentsusmkk 100606040208 Phpapp01Muaz AbdullahNo ratings yet
- Module 3Document3 pagesModule 3Charles JoseNo ratings yet
- CBTP Research PaperDocument67 pagesCBTP Research PaperEskindir Solomon100% (1)
- Vaginal Bleeding in Late PregnancyDocument139 pagesVaginal Bleeding in Late PregnancygibreilNo ratings yet
- The Importance of Nutrition & Early Stimulation for Brain DevelopmentDocument118 pagesThe Importance of Nutrition & Early Stimulation for Brain Developmentarie susantoNo ratings yet
- QuestionareDocument22 pagesQuestionareash aliNo ratings yet
- Implications of Early Pregnancy Obesity On Maternal, Fetal and Neonatal Health PDFDocument7 pagesImplications of Early Pregnancy Obesity On Maternal, Fetal and Neonatal Health PDFAndhika Dimas ANo ratings yet
- Epidemiology, Diagnosis, and Treatment of CerebrotendinousDocument11 pagesEpidemiology, Diagnosis, and Treatment of CerebrotendinousLuIz ZooZaNo ratings yet
- NCM 104 - M6 - DOH Programs Related To Family HealthDocument4 pagesNCM 104 - M6 - DOH Programs Related To Family HealthJollan Marie Buenvenida0% (1)
- 02 2002 EnglishDocument55 pages02 2002 EnglishAli ZeeshanNo ratings yet
- CounterclaimDocument2 pagesCounterclaimGessy Anne EstreraNo ratings yet
- Name: Demayo, Edson John C. Year & Section: BSN - 3A: Date: MARCH 16, 2021Document10 pagesName: Demayo, Edson John C. Year & Section: BSN - 3A: Date: MARCH 16, 2021Edson John DemayoNo ratings yet
- Abnormal Delivery Types and ManagementDocument12 pagesAbnormal Delivery Types and ManagementBadrul MunirNo ratings yet
- AOGDocument2 pagesAOGKing Nehpets AlczNo ratings yet
- Week 13 Care of Mother, ChildDocument27 pagesWeek 13 Care of Mother, ChildAnthonet AtractivoNo ratings yet
- Differentiate Between With AnswersDocument14 pagesDifferentiate Between With AnswersStephy SojanNo ratings yet
- Anemia FetalDocument14 pagesAnemia FetalyussyaNo ratings yet