You might also like
- Alakliphiles ProjDocument24 pagesAlakliphiles ProjReshmi Jadwani100% (2)
- A Study To Isolate The Alkaliphiles in The Mula Mutha River of Pune.Document3 pagesA Study To Isolate The Alkaliphiles in The Mula Mutha River of Pune.NikhilNo ratings yet
- AlkalophilesDocument10 pagesAlkalophilesrachnnaaNo ratings yet
- So Sánh Vsv Chịu MặnDocument7 pagesSo Sánh Vsv Chịu Mặndongthienb2No ratings yet
- Agricultural and Biological Chemistry: A A A A ADocument7 pagesAgricultural and Biological Chemistry: A A A A AAlfredo Gonzalez MateoNo ratings yet
- Extremophilic Microbes Alkaline PHDocument7 pagesExtremophilic Microbes Alkaline PHsashaikh1213No ratings yet
- Batch Studies On The Removal of Ni (II) From Aqueous Solution by Azolla FiliculoidesDocument5 pagesBatch Studies On The Removal of Ni (II) From Aqueous Solution by Azolla FiliculoidesalirezamdfNo ratings yet
- Harvesting of Wastewaters: Auto-Or Bioflocculation?: Scenedesmus Obliquus inDocument8 pagesHarvesting of Wastewaters: Auto-Or Bioflocculation?: Scenedesmus Obliquus inTeresa MataNo ratings yet
- Tetragenococcus Halophilus in Soy Sauce FermentationDocument6 pagesTetragenococcus Halophilus in Soy Sauce FermentationTâm Dương ĐứcNo ratings yet
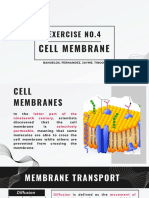
- Exercise No.4: Cell MembraneDocument28 pagesExercise No.4: Cell MembraneconnievelardeNo ratings yet
- Cowan 2021 Acidophiles Living in AcidDocument2 pagesCowan 2021 Acidophiles Living in AcidEllezar bakaNo ratings yet
- Systematic Characterization of Potential Cellulolytic Marine Actinobacteria Actinoalloteichus SP Mha15 PDFDocument7 pagesSystematic Characterization of Potential Cellulolytic Marine Actinobacteria Actinoalloteichus SP Mha15 PDFIT InventoryNo ratings yet
- Halophilic Enzymes5Document6 pagesHalophilic Enzymes5patel GayatriNo ratings yet
- Dermlim, Prasertsan, Doelle. 1999. Screening and Characterization of Bioflocculant Produced by Isolated Klebsiella SPDocument6 pagesDermlim, Prasertsan, Doelle. 1999. Screening and Characterization of Bioflocculant Produced by Isolated Klebsiella SPAzb 711No ratings yet
- Varsha Shivaji Kulkarni and Bhagyashree Shashikant KulkarniDocument11 pagesVarsha Shivaji Kulkarni and Bhagyashree Shashikant KulkarniCyriel Jade MendelloreNo ratings yet
- Partoza J.B Flowchart MethodologiesDocument4 pagesPartoza J.B Flowchart MethodologiesJenny PartozaNo ratings yet
- Fmicb 12 737244Document12 pagesFmicb 12 737244Natalija Atanasova-PancevskaNo ratings yet
- Escherichia Coli and Klebsiella: April 2011Document12 pagesEscherichia Coli and Klebsiella: April 2011ريام الموسويNo ratings yet
- Ethoxylates Alkylphenols PDFDocument15 pagesEthoxylates Alkylphenols PDFcausmicdanceNo ratings yet
- Ethoxylates Alkylphenols PDFDocument15 pagesEthoxylates Alkylphenols PDFcausmicdanceNo ratings yet
- Archae Bacteria Group 2 PresentationDocument14 pagesArchae Bacteria Group 2 PresentationDel-anne Laurice EstellinaNo ratings yet
- Chemical Carcinogenesis Handout 2023Document38 pagesChemical Carcinogenesis Handout 2023mpume lisahNo ratings yet
- The Acute Toxicity of Ammonia On Tilapia (Oreochromis Niloticus L.) Larvae and FingerlingsDocument7 pagesThe Acute Toxicity of Ammonia On Tilapia (Oreochromis Niloticus L.) Larvae and FingerlingsAbed ShareefNo ratings yet
- Inmovilizacion de CelulasDocument11 pagesInmovilizacion de CelulasFiorella Jennifer CZNo ratings yet
- Case Study 1-Stranger FishDocument2 pagesCase Study 1-Stranger FishNico RobinNo ratings yet
- Feed MixDocument4 pagesFeed MixTony1248No ratings yet
- Vinner e Pettersson 2011Document9 pagesVinner e Pettersson 2011Edson EscamillaNo ratings yet
- Alkaline PhosphataseDocument7 pagesAlkaline Phosphatasemostafa sheblNo ratings yet
- Elkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondDocument9 pagesElkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondAzb 711No ratings yet
- Segura 2014Document8 pagesSegura 2014dreyko 12No ratings yet
- Choi Et Al.. 1998. Characterization of An Extracellular Flocculating Substance Produced by A Planktonic Cyanobacterium, Anabaena SPDocument4 pagesChoi Et Al.. 1998. Characterization of An Extracellular Flocculating Substance Produced by A Planktonic Cyanobacterium, Anabaena SPAzb 711No ratings yet
- The Algae and Their General Characteristics: Susan E. DouglasDocument2 pagesThe Algae and Their General Characteristics: Susan E. DouglasLila MillerNo ratings yet
- Correlation Between Growth and Ergot Alkaloid BiosynthesisDocument4 pagesCorrelation Between Growth and Ergot Alkaloid BiosynthesisMonica Gaviria ArellanoNo ratings yet
- Ãvc Ãvî V3 VDocument6 pagesÃvc Ãvî V3 VSavannah Simone PetrachenkoNo ratings yet
- Jsir 61 (9) 690-704 PDFDocument15 pagesJsir 61 (9) 690-704 PDFAusteridad LopezNo ratings yet
- Ethoxylates AlkylphenolsDocument16 pagesEthoxylates AlkylphenolsBharatSubramonyNo ratings yet
- New Antiglycation and Enzyme Inhibitors From Parmotrema CooperiDocument7 pagesNew Antiglycation and Enzyme Inhibitors From Parmotrema CooperinoibinhyenchimhotNo ratings yet
- Clean Soil Air Water 2011Document9 pagesClean Soil Air Water 2011Rosmeri Eldy Nnhmn SllcNo ratings yet
- 128 2012 Article 598Document6 pages128 2012 Article 598PropsyMicDreamyNo ratings yet
- Metal Tolerance of Bioluminescent Bacteria Isolated From Marine OrganismsDocument8 pagesMetal Tolerance of Bioluminescent Bacteria Isolated From Marine OrganismsCyriel Jade MendelloreNo ratings yet
- Hackenberger2012 PDFDocument7 pagesHackenberger2012 PDFWahyu FathurrachmanNo ratings yet
- 13 Biotech Bioeng2004Document10 pages13 Biotech Bioeng2004Fiorella Jennifer CZNo ratings yet
- Studies On Extraction of Cyanobacterial Polysaccharides (Sacran) From and Its Structures and PropertiesDocument158 pagesStudies On Extraction of Cyanobacterial Polysaccharides (Sacran) From and Its Structures and PropertiesMoza Maria Iasmina (IASMY)No ratings yet
- 4 HainesDocument7 pages4 HainesFernandoNo ratings yet
- Adaptation of Spirogyra Insignis ChloropDocument7 pagesAdaptation of Spirogyra Insignis ChloropmanyadokdanielNo ratings yet
- Alkaloids: December 2010Document33 pagesAlkaloids: December 2010Jessica Asitimbay ZuritaNo ratings yet
- Microbial Alkaline ProteasesDocument34 pagesMicrobial Alkaline Proteasessalin MajumdarNo ratings yet
- Archaebacteria: BY Bibaswan Pritam SristyDocument25 pagesArchaebacteria: BY Bibaswan Pritam SristySRISTY AGARWALNo ratings yet
- Diversity of Microorganisms NotesDocument8 pagesDiversity of Microorganisms NotesFrances Hannah San LuisNo ratings yet
- Environmental MicrobiologyDocument40 pagesEnvironmental MicrobiologyKristell ValverdeNo ratings yet
- Natural Surfactants (Krister Holmberg) PDFDocument12 pagesNatural Surfactants (Krister Holmberg) PDFJohnyNo ratings yet
- Biochemistry Prelims: Topic 1: Introduction To BiochemistryDocument6 pagesBiochemistry Prelims: Topic 1: Introduction To BiochemistrycassseeeyyyNo ratings yet
- Vibrio and Related Organism: NotesDocument11 pagesVibrio and Related Organism: NotesSeyha L. AgriFoodNo ratings yet
- Deng Et Al.. 2003. Characteristics of A Bioflocculant Produced by Bacillus Mucilaginosus and Its Use in Starch Wastewater TreatmentDocument6 pagesDeng Et Al.. 2003. Characteristics of A Bioflocculant Produced by Bacillus Mucilaginosus and Its Use in Starch Wastewater TreatmentAzb 711No ratings yet
- Achal-Pan2011 Article CharacterizationOfUreaseAndCarDocument9 pagesAchal-Pan2011 Article CharacterizationOfUreaseAndCarIbrahim SalahudinNo ratings yet
- Dassarma 2006Document12 pagesDassarma 2006Mayty_AlarconNo ratings yet
- Archeae 1aDocument8 pagesArcheae 1aAnanya TomerNo ratings yet
- Hektoen Enteric Agar ProtocolDocument13 pagesHektoen Enteric Agar ProtocolYakisoba menNo ratings yet
- Tổng quan về vi khuẩn axit lacticDocument14 pagesTổng quan về vi khuẩn axit lacticMth RedNo ratings yet
- PLC Seminar 2 2022feb24Document1 pagePLC Seminar 2 2022feb24GhanshyamPatankarNo ratings yet
- ESD EXP5 Purva 202151002Document14 pagesESD EXP5 Purva 202151002GhanshyamPatankarNo ratings yet
- Experiment No. 3 - RIPDocument3 pagesExperiment No. 3 - RIPGhanshyamPatankarNo ratings yet
- ESD EXP5 Purva 202151002Document14 pagesESD EXP5 Purva 202151002GhanshyamPatankarNo ratings yet
- ESD EXP5 Purva 202151002Document14 pagesESD EXP5 Purva 202151002GhanshyamPatankarNo ratings yet
- SR - No. Name of Student Seminar Topic Seminar TopicDocument2 pagesSR - No. Name of Student Seminar Topic Seminar TopicGhanshyamPatankarNo ratings yet
- Name: Ghanshyam Patankar Date of Submission: 13-03-2021 ID: 202150015 Experiment No. 5Document8 pagesName: Ghanshyam Patankar Date of Submission: 13-03-2021 ID: 202150015 Experiment No. 5GhanshyamPatankarNo ratings yet
- Name: Ghanshyam Patankar Date of Submission: 03-03-2021 ID: 202150015 Experiment No. 3Document5 pagesName: Ghanshyam Patankar Date of Submission: 03-03-2021 ID: 202150015 Experiment No. 3GhanshyamPatankarNo ratings yet
- Wireless Channel Small-Scale Fading: 1/6/2021 Dr. Bharati Singh 1Document56 pagesWireless Channel Small-Scale Fading: 1/6/2021 Dr. Bharati Singh 1GhanshyamPatankarNo ratings yet
- HC20297293 PDFDocument20 pagesHC20297293 PDFGhanshyamPatankarNo ratings yet
- ADSP LAB, First Year M. Tech. E&TCDocument8 pagesADSP LAB, First Year M. Tech. E&TCGhanshyamPatankarNo ratings yet
- Wireless Channel Large Scale LossesDocument114 pagesWireless Channel Large Scale LossesGhanshyamPatankarNo ratings yet
- HC20297293 PDFDocument20 pagesHC20297293 PDFGhanshyamPatankarNo ratings yet
- Life Sci Final Question BankDocument3 pagesLife Sci Final Question BankGhanshyamPatankarNo ratings yet
- Uav Raspberry Pi Tutorial Without VideosDocument43 pagesUav Raspberry Pi Tutorial Without VideossNo ratings yet
- Roll No.:: Answer Sheet: Online ExaminationDocument1 pageRoll No.:: Answer Sheet: Online ExaminationGhanshyamPatankarNo ratings yet
- MMWC DPK WMC Bas Networking Lab ASG Rmip RGK Rmip RGK Mncc-I Rad Mncc-I RadDocument1 pageMMWC DPK WMC Bas Networking Lab ASG Rmip RGK Rmip RGK Mncc-I Rad Mncc-I RadGhanshyamPatankarNo ratings yet
- 09 03 2020 11.41.15Document1 page09 03 2020 11.41.15GhanshyamPatankarNo ratings yet
- Project Resarch Proposal Main 2Document27 pagesProject Resarch Proposal Main 2GhanshyamPatankar75% (4)
- Uav Raspberry Pi Tutorial Without VideosDocument1 pageUav Raspberry Pi Tutorial Without VideosGhanshyamPatankarNo ratings yet
- Aladin 2017Document6 pagesAladin 2017Dwi ermawatiNo ratings yet
- Phyton 27 MSDSDocument2 pagesPhyton 27 MSDSBernardo UenoNo ratings yet
- Biological ManagementDocument27 pagesBiological ManagementpatrickkayeNo ratings yet
- IGNTU Econtent 674613883400 B.SC EnvironmentalSciences 2 PallaviDas EnvironmentalPollutionandHumanHealth 2Document136 pagesIGNTU Econtent 674613883400 B.SC EnvironmentalSciences 2 PallaviDas EnvironmentalPollutionandHumanHealth 2Neeraj Raushan KanthNo ratings yet
- Fick Second LawDocument9 pagesFick Second LawJohnny WoodsNo ratings yet
- Fardis EC8-3 Member Models - 0Document75 pagesFardis EC8-3 Member Models - 0Wendirad BeshadaNo ratings yet
- Pharmacology Notes (Introduction To Pharmacology)Document16 pagesPharmacology Notes (Introduction To Pharmacology)BRYCE WILLIAM GONo ratings yet
- Redox EquilibriaDocument2 pagesRedox Equilibriafunkykid80No ratings yet
- Sarcosine MsdsDocument41 pagesSarcosine MsdsAnonymous ZVvGjtUGNo ratings yet
- Chapter Four, Cycloalkanes (Part One - Monocyclic Alkane)Document8 pagesChapter Four, Cycloalkanes (Part One - Monocyclic Alkane)Amin JamjahNo ratings yet
- pg1 PDFDocument1 pagepg1 PDFSanjeev GuptaNo ratings yet
- DELTA-FC 1022 Tds - EngDocument2 pagesDELTA-FC 1022 Tds - EngAhmed HassanNo ratings yet
- Microfine Cement Grouts & Applications in Grouting PracticeDocument38 pagesMicrofine Cement Grouts & Applications in Grouting PracticeGrigas GeorgeNo ratings yet
- Chem2 Lesson 1 - Kinetic Molecular Model of Liquids and SolidsDocument8 pagesChem2 Lesson 1 - Kinetic Molecular Model of Liquids and SolidsCarl EscalaNo ratings yet
- Pneumatic Auto Feed Drilling Machine With Indexing Machine: SynopsisDocument30 pagesPneumatic Auto Feed Drilling Machine With Indexing Machine: SynopsisMaruthi JacsNo ratings yet
- Part A: Multiple - Choice QuestionsDocument14 pagesPart A: Multiple - Choice QuestionsGora PostingNo ratings yet
- 409 Data BulletinDocument12 pages409 Data BulletinWilliam PaivaNo ratings yet
- Who We Are: Case Study-Mahindra ChlorisDocument25 pagesWho We Are: Case Study-Mahindra ChlorisSUNIDHI VERMANo ratings yet
- (2017) Toxicological Effects of Glycyrrhiza Glabra (Licorice) A ReviewDocument16 pages(2017) Toxicological Effects of Glycyrrhiza Glabra (Licorice) A ReviewicaNo ratings yet
- HEPA Cleanroom Filter Testing PDFDocument4 pagesHEPA Cleanroom Filter Testing PDFprovaoralNo ratings yet
- Dissolution TestDocument5 pagesDissolution TestAlecza Mae SavellaNo ratings yet
- 51314-3985-Methanol-Induced Internal Stress CorrosDocument18 pages51314-3985-Methanol-Induced Internal Stress CorrosMahmoud GamalNo ratings yet
- Module 2 - Fire SafetyDocument68 pagesModule 2 - Fire Safetyjairath100% (1)
- High Voltage Cable JointingDocument7 pagesHigh Voltage Cable Jointingscrapmail9No ratings yet
- Artificial Blood: Dr. Pragasam Viswanathan, Professor, SBSTDocument21 pagesArtificial Blood: Dr. Pragasam Viswanathan, Professor, SBSTMaru Mengesha Worku 18BBT0285No ratings yet
- Ray Bowl MillDocument9 pagesRay Bowl MillAnup MinjNo ratings yet
- What Pipeliners Need To Know About Induction Bends PDFDocument20 pagesWhat Pipeliners Need To Know About Induction Bends PDFmarcos2dami2o2de2azeNo ratings yet
- Ace Your Food Science Projects PDFDocument367 pagesAce Your Food Science Projects PDFashqNo ratings yet
- Tds Chemical Resistant CoatingDocument3 pagesTds Chemical Resistant CoatingUtilities2No ratings yet
- Salting-Out Crystallisation Using NH Ina Laboratory-Scale Gas Lift ReactorDocument10 pagesSalting-Out Crystallisation Using NH Ina Laboratory-Scale Gas Lift ReactorChester LowreyNo ratings yet