You might also like
- Homeostasis CrosswordDocument2 pagesHomeostasis CrosswordNirmala Josephine100% (1)
- Fast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlFrom EverandFast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlNo ratings yet
- Assignment 7 Hmpyc80Document16 pagesAssignment 7 Hmpyc80Tiffany SmithNo ratings yet
- Anatomy of Ear 1Document18 pagesAnatomy of Ear 1VikrantNo ratings yet
- Dehydroepiandrosterone Role PDFDocument8 pagesDehydroepiandrosterone Role PDFRiskaNo ratings yet
- Q 2002 Society of The European Journal of Endocrinology: Via Free AccessDocument10 pagesQ 2002 Society of The European Journal of Endocrinology: Via Free AccessThomas ColodelNo ratings yet
- The Evolving Role of Selenium in The Treatment of PDFDocument7 pagesThe Evolving Role of Selenium in The Treatment of PDFAkun TerorNo ratings yet
- Dehydroepiandrosterone (DHEA) : Hypes and HopesDocument13 pagesDehydroepiandrosterone (DHEA) : Hypes and HopesFernanda PérezNo ratings yet
- DRESS Syndrome A Detailed InsightDocument12 pagesDRESS Syndrome A Detailed InsightLeba SouloungerNo ratings yet
- International Immunopharmacology: SciencedirectDocument7 pagesInternational Immunopharmacology: SciencedirectPrabhjot SinghNo ratings yet
- Therapeutic Strategies in Psoriasis Patients With Psoriatic ArthritisDocument15 pagesTherapeutic Strategies in Psoriasis Patients With Psoriatic ArthritisCHONG WEI SHENGNo ratings yet
- 2019 Article 998 CompressedDocument11 pages2019 Article 998 CompressedAnonymous gzXHCYNo ratings yet
- 1 s2.0 S0161589019301592 MainDocument10 pages1 s2.0 S0161589019301592 Mainlaura quicenoNo ratings yet
- Asthma and Corticosteroids Time For A More Precise Approach To TreatmentDocument3 pagesAsthma and Corticosteroids Time For A More Precise Approach To TreatmentIfan SastraNo ratings yet
- Viral Infections in Allergy and Immunology: How Allergic Inflammation Influences Viral Infections and IllnessDocument12 pagesViral Infections in Allergy and Immunology: How Allergic Inflammation Influences Viral Infections and IllnessFajar SukmajayaNo ratings yet
- Hashimoto's ResearchDocument10 pagesHashimoto's Researchvgj6qr57c4No ratings yet
- Reyes-Pedia Journal PDFDocument8 pagesReyes-Pedia Journal PDFMicaNo ratings yet
- Update On Severe Asthma: What We Know and What We NeedDocument8 pagesUpdate On Severe Asthma: What We Know and What We Needtaksim2011No ratings yet
- DHEA Cancer EnvelhecimentoDocument10 pagesDHEA Cancer EnvelhecimentoDr. Ivan MateusNo ratings yet
- Kamei 1996Document8 pagesKamei 1996Dewi SariNo ratings yet
- Brandt 2020Document13 pagesBrandt 2020Hendri WahyudiNo ratings yet
- Meln Et Al-2012-Journal of Internal MedicineDocument13 pagesMeln Et Al-2012-Journal of Internal MedicineFadila PasyahNo ratings yet
- Hipotioridismo en Ratas Disminución de Energia y AyunoDocument11 pagesHipotioridismo en Ratas Disminución de Energia y AyunoPETERNo ratings yet
- Stress and ImmunityDocument5 pagesStress and ImmunitymadeNo ratings yet
- Pergamon: Glucocorticoids and TuberculosisDocument5 pagesPergamon: Glucocorticoids and TuberculosisGeorge AtaherNo ratings yet
- Anti IL 4 13 Based Therapy in AsthmaDocument5 pagesAnti IL 4 13 Based Therapy in AsthmaMoch Ridwan Pujiar PamungkasNo ratings yet
- Fphar-02-00050 Vitamin D and Susceptibility of Chronic Lung DiseasesDocument10 pagesFphar-02-00050 Vitamin D and Susceptibility of Chronic Lung DiseasesFiterman AdrianNo ratings yet
- Jurnal Alergi - Rendra PS - Phosphodiesterase 4 Inhibitors in Allergic Rhinitis - 125114Document16 pagesJurnal Alergi - Rendra PS - Phosphodiesterase 4 Inhibitors in Allergic Rhinitis - 125114Anggoro SatrioNo ratings yet
- Letter To The Editor: A, B C C C D e A ADocument3 pagesLetter To The Editor: A, B C C C D e A Amacan77No ratings yet
- Immunomodulators For AsthmaDocument7 pagesImmunomodulators For AsthmadarmariantoNo ratings yet
- Asma NewsDocument31 pagesAsma NewsEmmanuel HernandezNo ratings yet
- Asthma and COPD in Alpha 1 Antitrypsin Deficiency Evidence For The Dutch HypothesisDocument10 pagesAsthma and COPD in Alpha 1 Antitrypsin Deficiency Evidence For The Dutch HypothesisMohammed SaleetNo ratings yet
- Review Selenium and Endocrine Systems: and John R ArthurDocument11 pagesReview Selenium and Endocrine Systems: and John R ArthurcolaboradorforaneoNo ratings yet
- Concise Clinical Review: Role of Biologics in AsthmaDocument13 pagesConcise Clinical Review: Role of Biologics in AsthmaHossam HamzaNo ratings yet
- Dayan 2013Document12 pagesDayan 2013omar rodriguezNo ratings yet
- Cardona Et Al 2017 AllergyDocument8 pagesCardona Et Al 2017 AllergyAnasthasia hutagalungNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFAPOTEK DE DUA JAYA FARMANo ratings yet
- Gustavo H. Oliveira-Paula, Riccardo Lacchini, Fernanda B. Coeli-Lacchini, Heitor Moreno Junior, Jose E. Tanus-SantosDocument5 pagesGustavo H. Oliveira-Paula, Riccardo Lacchini, Fernanda B. Coeli-Lacchini, Heitor Moreno Junior, Jose E. Tanus-SantosdeddyNo ratings yet
- Dhea DescriptionDocument23 pagesDhea DescriptionNemes Kovács KataNo ratings yet
- Parasitic HelminthsDocument9 pagesParasitic HelminthsAnanta SwargiaryNo ratings yet
- Control AsthmaDocument3 pagesControl AsthmaAndi Suchy Qumala SarieNo ratings yet
- En 14Document8 pagesEn 14buo_souzaNo ratings yet
- New Insights Into The Relationship Between Airway Inflammation and AsthmaDocument11 pagesNew Insights Into The Relationship Between Airway Inflammation and Asthmaapi-3736350No ratings yet
- Ratna Et AlDocument6 pagesRatna Et AleditorijmrhsNo ratings yet
- Covid e AsmaDocument11 pagesCovid e AsmaIsabela ColenNo ratings yet
- Autoimmune Thyroid Disease Genes Identified in Non-CaucasiansDocument10 pagesAutoimmune Thyroid Disease Genes Identified in Non-CaucasiansHarnadi WonogiriNo ratings yet
- F - 2178 JCD Role of Apoptosis in Amplifying Inflammatory Responses in Lung Disease - PDF - 2981 PDFDocument13 pagesF - 2178 JCD Role of Apoptosis in Amplifying Inflammatory Responses in Lung Disease - PDF - 2981 PDFRanti WidiyantiNo ratings yet
- Diferentes Fases PDFDocument10 pagesDiferentes Fases PDFJudith Torres VillaNo ratings yet
- Wilson2006. Antihistamine For Asthma TherapyDocument10 pagesWilson2006. Antihistamine For Asthma TherapyMayke EviNo ratings yet
- Vitamin D and Thyroid Disease: To D or Not To D?: Mini ReviewDocument6 pagesVitamin D and Thyroid Disease: To D or Not To D?: Mini Reviewmammon mammonovNo ratings yet
- T-Helper Type 2-Driven Inflammation Defines Major. 2009Document8 pagesT-Helper Type 2-Driven Inflammation Defines Major. 2009mafe.z.solarte87No ratings yet
- Essay: Rethinking The Pathogenesis of AsthmaDocument6 pagesEssay: Rethinking The Pathogenesis of AsthmaConstanza Rada Von FurstenbergNo ratings yet
- Amiodarona y TiroidesDocument17 pagesAmiodarona y TiroidesTony Jose Bermejo ZuñigaNo ratings yet
- Bacterial Meningitis 19Document10 pagesBacterial Meningitis 19caliptra36No ratings yet
- Zhu 18 05Document8 pagesZhu 18 05ABNo ratings yet
- Paper Fiebre ReumDocument10 pagesPaper Fiebre ReumSara OchoaNo ratings yet
- Antihistamines and Asthma Patients 2002Document3 pagesAntihistamines and Asthma Patients 2002KhalidLabadiNo ratings yet
- JurnalDocument11 pagesJurnalLaksita Balqis MaharaniNo ratings yet
- Jbcahs 4 20Document3 pagesJbcahs 4 20Deana FrinsNo ratings yet
- J Aller 2019 12 011Document9 pagesJ Aller 2019 12 011Nelson MoralesNo ratings yet
- Hypertension: A New Treatment For An Old Disease? Targeting The Immune SystemDocument55 pagesHypertension: A New Treatment For An Old Disease? Targeting The Immune SystemgpatwallahNo ratings yet
- Acute DistressDocument15 pagesAcute Distresspablopg_5No ratings yet
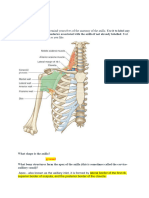
- The Human Body: An Orientation: © 2012 Pearson Education, IncDocument51 pagesThe Human Body: An Orientation: © 2012 Pearson Education, IncMohammad DweibNo ratings yet
- B UTS Bahasa Inggris Biologi 2020Document5 pagesB UTS Bahasa Inggris Biologi 2020kontol asuNo ratings yet
- (Answer Key 6.1) - (Life Processes)Document6 pages(Answer Key 6.1) - (Life Processes)Ashwin KNo ratings yet
- Revision of SpacesDocument6 pagesRevision of SpacesFrancesca vitaleNo ratings yet
- NEET Gold Biology Animal KingdomDocument52 pagesNEET Gold Biology Animal KingdomNagendra PrasadNo ratings yet
- Neuropsychology Notes 2Document10 pagesNeuropsychology Notes 2LiviaNo ratings yet
- IM 2B Long Quiz 2 Reviewer 3.2 MTDocument318 pagesIM 2B Long Quiz 2 Reviewer 3.2 MTSymon MillarNo ratings yet
- Persiapan Rikkes 2019: Instalasi Laboratorium Rumah Sakit Bhayangkara Polda D.I. YogyakartaDocument6 pagesPersiapan Rikkes 2019: Instalasi Laboratorium Rumah Sakit Bhayangkara Polda D.I. Yogyakartaferi rotinsuluNo ratings yet
- Osmosis and DiffusionDocument21 pagesOsmosis and DiffusionAtani ChineduNo ratings yet
- OpioidsDocument28 pagesOpioidsAmal MohammedNo ratings yet
- Exocrine Glands-Endocrine GlandsDocument8 pagesExocrine Glands-Endocrine GlandsneerleoNo ratings yet
- Circulatory System - Answer KeyDocument5 pagesCirculatory System - Answer Keyroblox dude ibbuzummerNo ratings yet
- 135 - Possible MecanismsDocument19 pages135 - Possible MecanismsEduardo BacelarNo ratings yet
- NewDocument93 pagesNewNarayan TiwariNo ratings yet
- Journal Song2014 FucoidanDocument6 pagesJournal Song2014 FucoidanGandhis Apri WidhayantiNo ratings yet
- Bobcat Compact Excavator E10 Service Manual 6986788Document22 pagesBobcat Compact Excavator E10 Service Manual 6986788andrewshelton250683psa98% (66)
- 7 - Isso Format QP - Set-2Document7 pages7 - Isso Format QP - Set-2pavanvanguriNo ratings yet
- JAP130 Slides 6Document5 pagesJAP130 Slides 6smpn1 PabuaranNo ratings yet
- Malignant Pleural Effusion and Its Current Management A ReviewDocument21 pagesMalignant Pleural Effusion and Its Current Management A ReviewJohn Sebastian ValenciaNo ratings yet
- TRT Tinnitus TreatmentDocument16 pagesTRT Tinnitus TreatmentKarina CastroNo ratings yet
- Brain SciencesDocument25 pagesBrain SciencesIratanFernandesNo ratings yet
- Embryo JohariDocument25 pagesEmbryo JohariTejas PLNo ratings yet
- Degradation of TG: 1 - Fat catabolism (lipolysis) 2.β-Oxidation of Fatty acids (fatty acid oxidation)Document26 pagesDegradation of TG: 1 - Fat catabolism (lipolysis) 2.β-Oxidation of Fatty acids (fatty acid oxidation)Aboubakar Moalim Mahad moh'dNo ratings yet
- Bio ReviewerDocument4 pagesBio ReviewerCaptain YooNo ratings yet
- Cal EcgDocument7 pagesCal EcgNur Amirah FarhanahNo ratings yet
- Life Processes 4Document18 pagesLife Processes 4Generic nameNo ratings yet
- ACT REAL BIOLOGY DR MAI ABD EL SALAM(1)Document30 pagesACT REAL BIOLOGY DR MAI ABD EL SALAM(1)ZeyadNo ratings yet