You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Jamaotolaryngology Eliezer 2020 Lo 200003Document2 pagesJamaotolaryngology Eliezer 2020 Lo 200003Pierre FlorentínNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Hydrogen Sulfide Toxicity Lactic Acid Inflammatory ProcessesDocument46 pagesHydrogen Sulfide Toxicity Lactic Acid Inflammatory ProcessesPierre FlorentínNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Ijcmr 1756 v1 PDFDocument4 pagesIjcmr 1756 v1 PDFAhmad KhairulNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Corona 1-S2.0-S2090123220300540-Main PDFDocument26 pagesCorona 1-S2.0-S2090123220300540-Main PDFshf.mxlikNo ratings yet
- DocumentDocument13 pagesDocumentPierre FlorentínNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Climate Risk ManagementDocument16 pagesClimate Risk ManagementPierre FlorentínNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Jems201707 DL PDFDocument70 pagesJems201707 DL PDFPierre FlorentínNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Renal Failure: Causes, Symptoms, and TreatmentDocument38 pagesRenal Failure: Causes, Symptoms, and TreatmentPierre FlorentínNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Lead Basics 1Document19 pagesLead Basics 1Steven D Owen Sr100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- SK716 Pengintegrasian Mekanisme Nilai Karbon (English)Document13 pagesSK716 Pengintegrasian Mekanisme Nilai Karbon (English)Isti HanifahNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Design Calculations of Thrust Blocks at Lower Indira: L&T Construction Water & Effluent Treatment ICDocument3 pagesDesign Calculations of Thrust Blocks at Lower Indira: L&T Construction Water & Effluent Treatment IClagnajit dasNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Memories of HomeDocument14 pagesMemories of HomeMary Francis Edmer SayconNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- 2008 Infosys Model QuestionsDocument23 pages2008 Infosys Model Questionsapi-3824713No ratings yet
- Midwifery All Year ProgramDocument3 pagesMidwifery All Year Programfiraol mokonnenNo ratings yet
- SR. NO. Capacity/Descirption of Machine Client: A.C. Generator StatorsDocument5 pagesSR. NO. Capacity/Descirption of Machine Client: A.C. Generator Statorsmtj4uNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- EU Food Label GuidanceDocument99 pagesEU Food Label GuidanceMunteanu CristianNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Life Time Prediction of GRP Piping SystemsDocument15 pagesLife Time Prediction of GRP Piping SystemsRamon FrenchNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Pasture FatteningDocument15 pagesPasture FatteningKartika PurnamasariNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Ras Abu Aboud Stadium Daily Report 040 (20190613)Document4 pagesRas Abu Aboud Stadium Daily Report 040 (20190613)tuan50% (2)
- Kyle 106 - 119 - 131 - 141Document1 pageKyle 106 - 119 - 131 - 141Gerson SouzaNo ratings yet
- Shock Classification and PathophysiologyDocument40 pagesShock Classification and PathophysiologyErick Anca100% (2)
- Unit 4 Early HumansDocument8 pagesUnit 4 Early HumansSepfira ReztikaNo ratings yet
- A Robust Firearm Identification Algorithm ProposalDocument7 pagesA Robust Firearm Identification Algorithm ProposalEuneel EscalaNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Lista Arancelaria EUADocument547 pagesLista Arancelaria EUAvilisbeth18No ratings yet
- Kyocera Servise ManualDocument613 pagesKyocera Servise ManualleukadiosNo ratings yet
- Bio Sem2 EssayDocument2 pagesBio Sem2 EssayEileen WongNo ratings yet
- Explore the beaches and parks of Labuan IslandDocument2 pagesExplore the beaches and parks of Labuan IslandDudeNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Resource Guide: Three Phase PowerDocument8 pagesResource Guide: Three Phase PowerddNo ratings yet
- Bio-Sil ULTRA 1050: Medically Pure Platinum-Cured Silicone Tubing With An Ultra-Smooth Inner SurfaceDocument2 pagesBio-Sil ULTRA 1050: Medically Pure Platinum-Cured Silicone Tubing With An Ultra-Smooth Inner SurfaceLari GrossiNo ratings yet
- Introduction To The Philosophy of The Human PersonDocument14 pagesIntroduction To The Philosophy of The Human PersonDan Niel50% (2)
- Deks Olje ArticleDocument5 pagesDeks Olje ArticleTomMacNaughtonNo ratings yet
- EE 102 Cabric Final Spring08 o Id15Document10 pagesEE 102 Cabric Final Spring08 o Id15Anonymous TbHpFLKNo ratings yet
- A History of The Jewish People in The Time of Jesus Christ (1891) Index Schürer, Emil, 1844-1910Document132 pagesA History of The Jewish People in The Time of Jesus Christ (1891) Index Schürer, Emil, 1844-1910David Bailey100% (2)
- Meralco Bill 330370940102 04142023Document2 pagesMeralco Bill 330370940102 04142023Jha CruzNo ratings yet
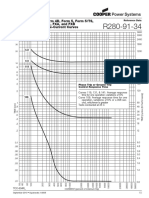
- Diagrama 4 AMBIENT AIR TEMPER..Document1 pageDiagrama 4 AMBIENT AIR TEMPER..Gustavo PérezNo ratings yet
- CD 0400 CH 4 X 100 ML: For in Vitro Diagnostic Use Only. LinearityDocument1 pageCD 0400 CH 4 X 100 ML: For in Vitro Diagnostic Use Only. LinearityNguyễn ThơiNo ratings yet
- The Image of The CityDocument17 pagesThe Image of The CityMay Rose ParagasNo ratings yet
- Musical Siren Project Report Under 40 CharactersDocument10 pagesMusical Siren Project Report Under 40 Charactersvinod kapateNo ratings yet
- 4 Floral ClockDocument4 pages4 Floral ClockmiguelibasterNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)