You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (843)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (346)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- First Records of Urban Invasive Cryptotermes BreviDocument5 pagesFirst Records of Urban Invasive Cryptotermes BrevinanchomartinNo ratings yet
- Behavioral and Electrophysiological Effects of AjoDocument15 pagesBehavioral and Electrophysiological Effects of AjonanchomartinNo ratings yet
- Draft Genomes of Amanita Jacksonii Cerat PDFDocument14 pagesDraft Genomes of Amanita Jacksonii Cerat PDFnanchomartinNo ratings yet
- Aftermath of Mountain Pine Beetle OutbreDocument19 pagesAftermath of Mountain Pine Beetle OutbrenanchomartinNo ratings yet
- Manual For Saving Water: Water Is A Diminishing Resource!Document5 pagesManual For Saving Water: Water Is A Diminishing Resource!nanchomartinNo ratings yet
- Bamboo Flooring Manufacturing Unit PDFDocument18 pagesBamboo Flooring Manufacturing Unit PDFRij GurungNo ratings yet
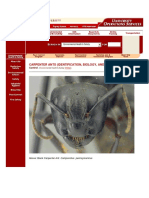
- Carpenter Ants (Identification, Biology, and Control)Document5 pagesCarpenter Ants (Identification, Biology, and Control)ViolintsarNo ratings yet
- CD Township Complaint OldDocument6 pagesCD Township Complaint Oldbalram4uNo ratings yet
- ShrublandsDocument2 pagesShrublandseristretto04No ratings yet
- Class 10Document2 pagesClass 10krishnithin19No ratings yet
- Example of Research Paper About Illegal LoggingDocument4 pagesExample of Research Paper About Illegal Loggingsuz1sezibys2100% (1)
- Test Conny ReadingDocument10 pagesTest Conny ReadingAndres VerdugoNo ratings yet
- 02teamwork2 Unit5 L2Document7 pages02teamwork2 Unit5 L2marcosgarciacarreraNo ratings yet
- Forests IgcseDocument21 pagesForests Igcseleenamuneeb1983No ratings yet
- M SC Forestry Syllabus (New Revised)Document53 pagesM SC Forestry Syllabus (New Revised)Vishal VijayanNo ratings yet
- FSC STD 50 001 - V1 2Document28 pagesFSC STD 50 001 - V1 2Marcelo Pérez VivancoNo ratings yet
- Brgy - ResolutionDocument2 pagesBrgy - ResolutionJohn Michael Austria PanganNo ratings yet
- Activity 3 Lets CheckDocument6 pagesActivity 3 Lets CheckDiana Mae Barcoma100% (13)
- Indonesian Fauna: ExampleDocument1 pageIndonesian Fauna: ExamplePradhita YudhaNo ratings yet
- Environment Quiz - 4Document5 pagesEnvironment Quiz - 4ITTeamNo ratings yet
- The GMs Real-World ReferenceDocument221 pagesThe GMs Real-World ReferenceAieie100% (1)
- Phenology Lab ReportDocument5 pagesPhenology Lab Reportapi-490258085No ratings yet
- Ventana Wilderness Watch: Kozy Kove Litigation UpdateDocument4 pagesVentana Wilderness Watch: Kozy Kove Litigation UpdateXatziliasNikosNo ratings yet
- Maps For Erebor AdventuresDocument2 pagesMaps For Erebor AdventuresI.M. AwesomemanNo ratings yet
- Field Guide To Florida Mangroves: Avicennia Laguncularia Conocarpus RhizophoraDocument2 pagesField Guide To Florida Mangroves: Avicennia Laguncularia Conocarpus RhizophoraXu ZhangNo ratings yet
- Abelhas Do RS - Wittmann 1990Document30 pagesAbelhas Do RS - Wittmann 1990NinaNo ratings yet
- Wellington Firewood Collection Areas For 2023Document2 pagesWellington Firewood Collection Areas For 2023tylerbottegaNo ratings yet
- Journal Entry 5.2.docx - Google Tài liệuDocument3 pagesJournal Entry 5.2.docx - Google Tài liệuBenNo ratings yet
- Ge 15 Reviewer 1 3Document5 pagesGe 15 Reviewer 1 3LORRAINE ANNE LAURENTENo ratings yet
- Telephone Chart As On 27 09 2022Document1 pageTelephone Chart As On 27 09 2022Save EnvironmentNo ratings yet
- SOP PlantationsDocument123 pagesSOP PlantationsDarnell SutherlandNo ratings yet
- PDF 20220220 164630 0000Document11 pagesPDF 20220220 164630 0000Devananda MeladathNo ratings yet
- Article - What Is A Keystone SpeciesDocument2 pagesArticle - What Is A Keystone SpeciesBob JoeNo ratings yet
- Geog Notes Summary PDFDocument20 pagesGeog Notes Summary PDFBaryaNo ratings yet
- On BiomesDocument11 pagesOn BiomesJM MejiaNo ratings yet