You might also like
- Marine Impacts of Seawater Desalination: Science, Management, and PolicyFrom EverandMarine Impacts of Seawater Desalination: Science, Management, and PolicyNo ratings yet
- Znachoretal 2015 HydrobiologiaDocument14 pagesZnachoretal 2015 HydrobiologiaAnindia MawaddatiNo ratings yet
- Macheretal 2016printedpaperDocument12 pagesMacheretal 2016printedpaperSixto Gutiérrez SaavedraNo ratings yet
- Fish Models in ToxicologyDocument13 pagesFish Models in ToxicologyDavid James PNo ratings yet
- Monoraphid and Naviculoid Diatoms from the Coastal Laurentian Great LakesFrom EverandMonoraphid and Naviculoid Diatoms from the Coastal Laurentian Great LakesAndrzej WitkowskiNo ratings yet
- Cell-Associated Water: Proceedings of a Workshop on Cell-Associated Water Held in Boston, Massachusetts, September, 1976From EverandCell-Associated Water: Proceedings of a Workshop on Cell-Associated Water Held in Boston, Massachusetts, September, 1976W. Drost-HansenNo ratings yet
- Advances in Phytoplankton Ecology: Applications of Emerging TechnologiesFrom EverandAdvances in Phytoplankton Ecology: Applications of Emerging TechnologiesNo ratings yet
- Regulation of Parasite PopulationsFrom EverandRegulation of Parasite PopulationsGerald EschNo ratings yet
- Kermarrec Et Al. 14, A NGS Approach To River Biomonitoring Using Benthic Diatoms. FSDocument16 pagesKermarrec Et Al. 14, A NGS Approach To River Biomonitoring Using Benthic Diatoms. FSjasminajacaNo ratings yet
- A Systematic Review of Marine-Based Species DistriDocument12 pagesA Systematic Review of Marine-Based Species DistriMaria Rosalie NARITNo ratings yet
- Robinsonetal 2011Document15 pagesRobinsonetal 2011roraysideblancoNo ratings yet
- Review Paper Measurement of Pollutant Toxicity To Fish I. Bioassay Methods For Acute ToxicityDocument29 pagesReview Paper Measurement of Pollutant Toxicity To Fish I. Bioassay Methods For Acute ToxicityLuciane ReisNo ratings yet
- Eustasy, High-Frequency Sea Level Cycles and Habitat HeterogeneityFrom EverandEustasy, High-Frequency Sea Level Cycles and Habitat HeterogeneityNo ratings yet
- Salamat Zarei 2014Document7 pagesSalamat Zarei 2014Jayanthi TNo ratings yet
- Molecular Basis of Biological Degradative processesFrom EverandMolecular Basis of Biological Degradative processesRichard BerlinNo ratings yet
- Kroeker 2013 Global Change BiologyDocument14 pagesKroeker 2013 Global Change Biologymary balansagNo ratings yet
- Toxicity Testing, Aquatic: M Sobanska, R Cesnaitis, and B Versonnen, European Chemicals Agency, Helsinki, FinlandDocument5 pagesToxicity Testing, Aquatic: M Sobanska, R Cesnaitis, and B Versonnen, European Chemicals Agency, Helsinki, FinlandMarcos AlbertoNo ratings yet
- 1 - Crespo-Medina - Environmental Controls On Microbial ComunitiesDocument10 pages1 - Crespo-Medina - Environmental Controls On Microbial ComunitiesLUCIA BEATRICE NECHIFOR GRIGORENo ratings yet
- Rotifers As Models in Toxicity Screening of Chemicals and Environmental SamplesDocument45 pagesRotifers As Models in Toxicity Screening of Chemicals and Environmental SamplesroraysideblancoNo ratings yet
- Use of The Zebrafish Embryo Toxicity Test For Risk Assessment Purpose Case StudyDocument12 pagesUse of The Zebrafish Embryo Toxicity Test For Risk Assessment Purpose Case StudyLouhana Moreira ReboucasNo ratings yet
- Microbial Community Analysis of Swine Wastewater Anaerobic Laggons by Next-Generation DNA SequencingDocument8 pagesMicrobial Community Analysis of Swine Wastewater Anaerobic Laggons by Next-Generation DNA SequencingDoulalas GiorgosNo ratings yet
- Contaminants of Emerging Concern in A Large Temperate EstuaryDocument14 pagesContaminants of Emerging Concern in A Large Temperate EstuaryAnonymous 7RI4DUksNo ratings yet
- Progress in Biophysics and Biophysical Chemistry: Volume 12From EverandProgress in Biophysics and Biophysical Chemistry: Volume 12J. A. V. ButlerNo ratings yet
- PHD Thesis On ZooplanktonDocument8 pagesPHD Thesis On Zooplanktonannameleknorthlasvegas100% (2)
- Gross Et Al., 2012 Chapter - Allelochemical Interactions Among Aquatic Primary ProducersDocument15 pagesGross Et Al., 2012 Chapter - Allelochemical Interactions Among Aquatic Primary Producersmanuela.passosNo ratings yet
- 2005 Volume 96 Marine Biotechnology IDocument261 pages2005 Volume 96 Marine Biotechnology IabavoNo ratings yet
- Fmars 2017 00107Document14 pagesFmars 2017 00107izzy officialNo ratings yet
- Advancing Environmental Toxicology Through Chemical Dosimetry: External Exposures Versus Tissue ResiduesDocument22 pagesAdvancing Environmental Toxicology Through Chemical Dosimetry: External Exposures Versus Tissue ResiduesLuis RamosNo ratings yet
- MBI 2020 First EtalDocument19 pagesMBI 2020 First Etalfernandosirgo1No ratings yet
- An Integrated Mark-Recapture and Genetic Approach To Estimate The Population Size of White Shark in South AfricaDocument14 pagesAn Integrated Mark-Recapture and Genetic Approach To Estimate The Population Size of White Shark in South AfricaAlexandra LvNo ratings yet
- Biomagnification in Marine Systems: The Perspective of An EcologistDocument7 pagesBiomagnification in Marine Systems: The Perspective of An EcologistAdafabsNo ratings yet
- Intracellular TransportFrom EverandIntracellular TransportKatherine Brehme WarrenNo ratings yet
- Flocculation in Natural and Engineered Environmental SystemsDocument438 pagesFlocculation in Natural and Engineered Environmental SystemsPaulaGuerraPintoNo ratings yet
- Impacts of The Deepwater Horizon Oil Spill Evaluated Using An End-To-End Ecosystem ModelDocument21 pagesImpacts of The Deepwater Horizon Oil Spill Evaluated Using An End-To-End Ecosystem ModelMichael SalenussaNo ratings yet
- Publications Phyto List1Document93 pagesPublications Phyto List1Hendry WijayantiNo ratings yet
- Marine Paleobiodiversity: Responses to Sea Level Cycles and PerturbationsFrom EverandMarine Paleobiodiversity: Responses to Sea Level Cycles and PerturbationsNo ratings yet
- Genetic Diversity of Total, Active and Culturable Marine Bacteria in Coastal SeawaterDocument11 pagesGenetic Diversity of Total, Active and Culturable Marine Bacteria in Coastal SeawaterMelody ChristineNo ratings yet
- Natural Product Reports (2014), 31 (11), 1510-1553. ,: Scifinder®Document14 pagesNatural Product Reports (2014), 31 (11), 1510-1553. ,: Scifinder®Nicolas Ignacio Aliaga MachucaNo ratings yet
- (John Townend) Practical Statistics For EnvironmenDocument2 pages(John Townend) Practical Statistics For Environmendouglas gitongaNo ratings yet
- Barnes - Review Oceanography and Marine BiologyDocument425 pagesBarnes - Review Oceanography and Marine BiologyKhalid al-ghifariNo ratings yet
- Comparing Immobilization Recovery and Stress IndicDocument11 pagesComparing Immobilization Recovery and Stress IndicMiguel Ángel Gallego DomínguezNo ratings yet
- PDFDocument485 pagesPDFOki NurpatriaNo ratings yet
- An Assessment of The MTR Aquatic Plant BDocument7 pagesAn Assessment of The MTR Aquatic Plant BDragana Vukov VučkovićNo ratings yet
- DNA Barcoding and Metabarcoding of Standardized Samples Reveal Patterns of Marine Benthic DiversityDocument17 pagesDNA Barcoding and Metabarcoding of Standardized Samples Reveal Patterns of Marine Benthic DiversityJennifer SilvaNo ratings yet
- FishesDocument17 pagesFishesEduardo MiguelesNo ratings yet
- (2010) Behavioural Ecotoxicology, An "Early Warning" Signal To Assess Environmental Quality - Hellou, J.Document12 pages(2010) Behavioural Ecotoxicology, An "Early Warning" Signal To Assess Environmental Quality - Hellou, J.Jerika Marie ArceoNo ratings yet
- AntLee 2013 EndophyteDocument11 pagesAntLee 2013 EndophytewiroNo ratings yet
- Chiu Etal 2020 J. BiogeogDocument12 pagesChiu Etal 2020 J. BiogeogLewis JonesNo ratings yet
- Pandey 2018Document12 pagesPandey 2018Luciane ReisNo ratings yet
- Mcginty 2018 Meps m601p109Document19 pagesMcginty 2018 Meps m601p109Mar LunaNo ratings yet
- Fish Physiology: Organic Chemical Toxicology of FishesFrom EverandFish Physiology: Organic Chemical Toxicology of FishesNo ratings yet
- DNA Barcoding As A Tool For Coral Reef ConservatioDocument14 pagesDNA Barcoding As A Tool For Coral Reef ConservatioRhaiza PabelloNo ratings yet
- SMWW 10010Document2 pagesSMWW 10010ALVARONo ratings yet
- 1 s2.0 S2214714415300040 MainDocument9 pages1 s2.0 S2214714415300040 MainEdwin ShajuNo ratings yet
- Xbox One X Manual en Da NL Fi FR de It No PT Es SV - EuDocument100 pagesXbox One X Manual en Da NL Fi FR de It No PT Es SV - EuMiguel Ángel Gallego DomínguezNo ratings yet
- Xbox One X Manual en Da NL Fi FR de It No PT Es SV - EuDocument100 pagesXbox One X Manual en Da NL Fi FR de It No PT Es SV - EuMiguel Ángel Gallego DomínguezNo ratings yet
- Shayegan 2016Document10 pagesShayegan 2016Miguel Ángel Gallego DomínguezNo ratings yet
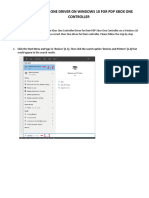
- Xbox One Driver Installation Guide For Windows 10Document4 pagesXbox One Driver Installation Guide For Windows 10Miguel Ángel Gallego DomínguezNo ratings yet
- PHD Thesis Guillaume Drillet PDFDocument170 pagesPHD Thesis Guillaume Drillet PDFMiguel Ángel Gallego DomínguezNo ratings yet
- Xbox One Driver Installation Guide For Windows 10Document4 pagesXbox One Driver Installation Guide For Windows 10Miguel Ángel Gallego DomínguezNo ratings yet
- PHD Thesis Guillaume Drillet PDFDocument170 pagesPHD Thesis Guillaume Drillet PDFMiguel Ángel Gallego DomínguezNo ratings yet
- Egg Production and Hatching Success of Four Acartia Species Under Different Temperature and Salinity Regimes Ernestina Castro-LongoriaDocument11 pagesEgg Production and Hatching Success of Four Acartia Species Under Different Temperature and Salinity Regimes Ernestina Castro-LongoriaMiguel Ángel Gallego DomínguezNo ratings yet
- Efectos Salinidad, Foto y Densidad en AcartiaDocument10 pagesEfectos Salinidad, Foto y Densidad en AcartiaMiguel Ángel Gallego DomínguezNo ratings yet
- Vibrio Spp. Control at Brine Shrimp, Artemia, Hatching: and EnrichmentDocument10 pagesVibrio Spp. Control at Brine Shrimp, Artemia, Hatching: and EnrichmentMiguel Ángel Gallego DomínguezNo ratings yet
- Holste Peck 2006Document11 pagesHolste Peck 2006Miguel Ángel Gallego DomínguezNo ratings yet
- Auratus Hybrid Striped Bass, Morone Hybrid Saxatilis M. Chrysops and Ocellated River Stingray, Potamotrygon MotoroDocument7 pagesAuratus Hybrid Striped Bass, Morone Hybrid Saxatilis M. Chrysops and Ocellated River Stingray, Potamotrygon MotoroMiguel Ángel Gallego DomínguezNo ratings yet
- On The Electroimmobilization of Fishes For Research and Practice: Opportunities, Challenges, and Research NeedsDocument34 pagesOn The Electroimmobilization of Fishes For Research and Practice: Opportunities, Challenges, and Research NeedsMiguel Ángel Gallego DomínguezNo ratings yet
- Comparing Immobilization Recovery and Stress IndicDocument11 pagesComparing Immobilization Recovery and Stress IndicMiguel Ángel Gallego DomínguezNo ratings yet
- Captureand Transportof Elasmobranchswithemphasisonthegreynurseshark CarchariastaurusDocument16 pagesCaptureand Transportof Elasmobranchswithemphasisonthegreynurseshark CarchariastaurusMiguel Ángel Gallego DomínguezNo ratings yet
- Chapter 21 - Anesthesia and Restraint of Laboratory FishDocument16 pagesChapter 21 - Anesthesia and Restraint of Laboratory FishMiguel Ángel Gallego DomínguezNo ratings yet
- HW 5, 448Document16 pagesHW 5, 448pdrogos02100% (1)
- IOS Developer - ExperiencedDocument37 pagesIOS Developer - Experiencedswornavidhya.mahadevanNo ratings yet
- Nitrosyl Complexes of RutheniumDocument26 pagesNitrosyl Complexes of RutheniumBenjamín Marc Ridgway de SassouNo ratings yet
- Holmium Laser - Fibers - Brochure - A4 - PB-006360 - Rev K - WebDocument4 pagesHolmium Laser - Fibers - Brochure - A4 - PB-006360 - Rev K - WebTigherMahNo ratings yet
- Handout 3 TMR4305 Lecture 13 09 2007Document5 pagesHandout 3 TMR4305 Lecture 13 09 2007sujaydsouza1987No ratings yet
- The Chemistry Maths Book: Erich SteinerDocument16 pagesThe Chemistry Maths Book: Erich SteinerDewi MariyamNo ratings yet
- Inorganic Chemistry 2 Main Exam (3) and MemoDocument11 pagesInorganic Chemistry 2 Main Exam (3) and MemoKgasu MosaNo ratings yet
- Fasteners For Use With PC BoardsDocument25 pagesFasteners For Use With PC BoardsJustin PhanNo ratings yet
- Surveying 2 Final ExamDocument3 pagesSurveying 2 Final ExamRodin James GabrilloNo ratings yet
- Dental CementDocument74 pagesDental CementAisha samreenNo ratings yet
- Serto Up To Date 33Document7 pagesSerto Up To Date 33Teesing BVNo ratings yet
- Generalized Digital Butterworth Filter Design: IEEE Transactions On Signal Processing May 1998Document31 pagesGeneralized Digital Butterworth Filter Design: IEEE Transactions On Signal Processing May 1998domnuljanNo ratings yet
- CS 515 Data Warehousing and Data MiningDocument5 pagesCS 515 Data Warehousing and Data MiningRahumal SherinNo ratings yet
- 21 10 26 Tastytrade ResearchDocument7 pages21 10 26 Tastytrade ResearchCSNo ratings yet
- Condensatoare MLCC-1837944Document130 pagesCondensatoare MLCC-1837944gigiNo ratings yet
- Lehmann Et Al 2013 PDFDocument13 pagesLehmann Et Al 2013 PDFJack HamiltonNo ratings yet
- ET4117 Electrical Machines and Drives Lecture5Document31 pagesET4117 Electrical Machines and Drives Lecture5farhan beighNo ratings yet
- Lecture 2 - Inverse Laplace TransformsDocument3 pagesLecture 2 - Inverse Laplace TransformsAmirah MapandiNo ratings yet
- Matlab Manual1Document25 pagesMatlab Manual1Richard BrooksNo ratings yet
- Lab Report Welding (SMAW) - FizaDocument13 pagesLab Report Welding (SMAW) - FizaCheng Yee71% (7)
- Aptitude Questions and Answers Sample Test 1Document11 pagesAptitude Questions and Answers Sample Test 1Udaya Kumar100% (1)
- Created by Ankur Sharma, Gurudayal Srivastava, Vikram Singh, Sanket TamhaneDocument12 pagesCreated by Ankur Sharma, Gurudayal Srivastava, Vikram Singh, Sanket TamhaneSankash SoodNo ratings yet
- 0w 30 Yağ ÖzellikleriDocument2 pages0w 30 Yağ ÖzellikleriahmetNo ratings yet
- Activities InterventionDocument2 pagesActivities InterventionDonabel Villasaya CariosNo ratings yet
- FOCSWATER.C Water Cooled Chiller - R134a - 2012!01!01Document15 pagesFOCSWATER.C Water Cooled Chiller - R134a - 2012!01!01ntt_121987No ratings yet
- S Bms ManualDocument44 pagesS Bms ManualnathanaelorNo ratings yet
- Compiled Blue Print and Syllabus XI HY 2022Document6 pagesCompiled Blue Print and Syllabus XI HY 2022shrutiNo ratings yet
- s10763 017 9799 1 PDFDocument19 pagess10763 017 9799 1 PDFJoy PascoNo ratings yet
- Synchronous Generators - 2 Marks Questions and AnswersDocument3 pagesSynchronous Generators - 2 Marks Questions and AnswersJoseph Harindranath67% (3)
- Examples S-Convex FunctionsDocument7 pagesExamples S-Convex Functionstradutora100% (2)