You might also like
- Xylitol Production From in Low-Cost Fermentation Media: Eucalyptus Wood HydrolysatesDocument7 pagesXylitol Production From in Low-Cost Fermentation Media: Eucalyptus Wood HydrolysatesJISENNIA MARLENY VARA GARCIANo ratings yet
- Acid Hydrolysis of Wheat Straw: A Kinetic StudyDocument10 pagesAcid Hydrolysis of Wheat Straw: A Kinetic StudyRicco Riyan KurniawanNo ratings yet
- 7 PBDocument7 pages7 PBat6315763No ratings yet
- Renewable Energy: I. Egüés, L. Serrano, D. Amendola, D.M. de Faveri, G. Spigno, J. LabidiDocument6 pagesRenewable Energy: I. Egüés, L. Serrano, D. Amendola, D.M. de Faveri, G. Spigno, J. LabidiRoshan GillNo ratings yet
- Cas Sales 2011Document9 pagesCas Sales 2011Camila NevesNo ratings yet
- Research 6Document7 pagesResearch 6UgoStanNo ratings yet
- Enzymatic Saccharification and Fermentation of CelDocument10 pagesEnzymatic Saccharification and Fermentation of CelSharmaNo ratings yet
- Integrated Production of Biofuels and Succinic Acid From Biomass After Thermochemical PretreatmentsDocument16 pagesIntegrated Production of Biofuels and Succinic Acid From Biomass After Thermochemical PretreatmentsJames TomosoriNo ratings yet
- ViewFreeArticle PDFDocument5 pagesViewFreeArticle PDFMaria Alyssa EdañoNo ratings yet
- EVALUATION OF SORGHUM STRAW HEMICELLULOSIC HYDROLYSATE FOR BIOTECHNOLOGICAL PRODUCTION OF XYLITOL BY Candida GuilliermondiiDocument6 pagesEVALUATION OF SORGHUM STRAW HEMICELLULOSIC HYDROLYSATE FOR BIOTECHNOLOGICAL PRODUCTION OF XYLITOL BY Candida Guilliermondiiprivaz81No ratings yet
- Fermentation of Sun Ower Seed Hull Hydrolysate To Ethanol by Pichia StipitisDocument8 pagesFermentation of Sun Ower Seed Hull Hydrolysate To Ethanol by Pichia StipitisJulio MoralesNo ratings yet
- BioRes 07 2 1540 Ding HO Compar Pretreat Strat Coconut Husk Sugars 2491 PDFDocument8 pagesBioRes 07 2 1540 Ding HO Compar Pretreat Strat Coconut Husk Sugars 2491 PDFNatasha Mgt JoharNo ratings yet
- Production of Ethanol From Soybean Hull Hydrolysate by Osmotolerant Candida Guilliermondii NRRL Y-2075Document7 pagesProduction of Ethanol From Soybean Hull Hydrolysate by Osmotolerant Candida Guilliermondii NRRL Y-2075Camila NevesNo ratings yet
- Production of Ethanol From The Hemicellulosic Fraction of Sunflower Meal BiomassDocument7 pagesProduction of Ethanol From The Hemicellulosic Fraction of Sunflower Meal BiomassJulio MoralesNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- Production of Fermentable Sugars by Combined Chemo-Enzymatic Hydrolysis of Cellulosic Material For Bioethanol ProductionDocument9 pagesProduction of Fermentable Sugars by Combined Chemo-Enzymatic Hydrolysis of Cellulosic Material For Bioethanol ProductionJerome JavierNo ratings yet
- °c/15psi) - Under This Condition, The Cellulose Content of Wheat Straw Was Increased byDocument9 pages°c/15psi) - Under This Condition, The Cellulose Content of Wheat Straw Was Increased byHet ThankiNo ratings yet
- Acid Hydrolysis of Sugarcane Bagasse For Lactic Acid ProductionDocument8 pagesAcid Hydrolysis of Sugarcane Bagasse For Lactic Acid ProductionAndressaGonçalvesNo ratings yet
- Ethanol CalculationDocument14 pagesEthanol CalculationMuhammad Azim Khan NiaziNo ratings yet
- Cannabis BiomassDocument8 pagesCannabis BiomassmarianaNo ratings yet
- Bioresources.: Bioconversion of Bamboo To Bioethanol Using The Two-Stage Organosolv and Alkali PretreatmentDocument9 pagesBioresources.: Bioconversion of Bamboo To Bioethanol Using The Two-Stage Organosolv and Alkali Pretreatmentcandra ndraNo ratings yet
- Bioteknologi Pangan-Dina 2018Document9 pagesBioteknologi Pangan-Dina 2018achn_tyaNo ratings yet
- 18 Teodor VintilaDocument7 pages18 Teodor VintilaleaderitNo ratings yet
- Vol2no9 8Document8 pagesVol2no9 8Peter Reinir DaadaNo ratings yet
- Amezcua-Allieri Et Al 2017 Study of Chemical and Enzymatic Hydrolysis of CellulosicDocument11 pagesAmezcua-Allieri Et Al 2017 Study of Chemical and Enzymatic Hydrolysis of CellulosicKike CamNo ratings yet
- Research Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionDocument10 pagesResearch Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionShankar AcharNo ratings yet
- Camargo 2014Document8 pagesCamargo 2014Julio MoralesNo ratings yet
- Effect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisDocument13 pagesEffect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisProAkademiaNo ratings yet
- Cellulose Extraction From Palm Kernel Cake Using Liquid Phase OxidationDocument12 pagesCellulose Extraction From Palm Kernel Cake Using Liquid Phase OxidationRidwan SantosoNo ratings yet
- Process Biochemistry: SciencedirectDocument9 pagesProcess Biochemistry: SciencedirectGayatri VishwaNo ratings yet
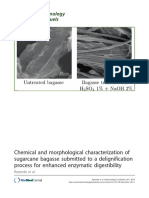
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Biodetoxification Amorphotheca Zhang Bao 2010Document15 pagesBiodetoxification Amorphotheca Zhang Bao 2010Anton van PuttenNo ratings yet
- Obtaining of Reducing Sugars From Kikuyu Grass (Pennisetum Clandestinum)Document4 pagesObtaining of Reducing Sugars From Kikuyu Grass (Pennisetum Clandestinum)Víctor Julio Daza MNo ratings yet
- Waste Management: K. Ziemin Ski, I. Romanowska, M. KowalskaDocument7 pagesWaste Management: K. Ziemin Ski, I. Romanowska, M. KowalskaMayra UlínNo ratings yet
- Ethanol Corn Cob - Phế Phụ LiệuDocument6 pagesEthanol Corn Cob - Phế Phụ LiệuDuy Tân NguyễnNo ratings yet
- Bioethanol Production From Banana Peels Using Different PretreatmentsDocument6 pagesBioethanol Production From Banana Peels Using Different PretreatmentsabdulhamidNo ratings yet
- Cellulolysis - Ethanol Production: Cellulolysis Is The Process of Breaking Down Cellulose IntoDocument9 pagesCellulolysis - Ethanol Production: Cellulolysis Is The Process of Breaking Down Cellulose Intoanushri birlaNo ratings yet
- Biohydrogen Production From Palm Oil Mill Effluent Pretreated by Chemical Methods Using Thermoanaerobacterium-Rich SludgeDocument8 pagesBiohydrogen Production From Palm Oil Mill Effluent Pretreated by Chemical Methods Using Thermoanaerobacterium-Rich SludgefransiscaNo ratings yet
- Nitric-Acid Hydrolysis of Miscanthus Giganteus To Sugars Fermented To BioethanolDocument2 pagesNitric-Acid Hydrolysis of Miscanthus Giganteus To Sugars Fermented To BioethanolMuhammadFakhrizalFahmiNo ratings yet
- Pineapple Peel Waste For Bioethanol ProductionDocument1 pagePineapple Peel Waste For Bioethanol ProductionAnge Cantor-dela CruzNo ratings yet
- Hydrolyse AcideDocument9 pagesHydrolyse AcideMohamed NaimiNo ratings yet
- Solid and Liquid Pineapple Waste Utilization For Lactic Acid Fermentation USING Lactobacillus DelbrueckiiDocument3 pagesSolid and Liquid Pineapple Waste Utilization For Lactic Acid Fermentation USING Lactobacillus Delbrueckiipatrica_23velezNo ratings yet
- Pretreatment Conditions of Rice Straw For Simultaneous Hydrogen and Ethanol Fermentation by Mixed CultureDocument8 pagesPretreatment Conditions of Rice Straw For Simultaneous Hydrogen and Ethanol Fermentation by Mixed CultureAldrienNo ratings yet
- Production of Acetone-Butanol-ethanol From Spoilage Date PalmDocument7 pagesProduction of Acetone-Butanol-ethanol From Spoilage Date PalmI-Twins RgnNo ratings yet
- Improved Upstream Processing For Detoxification and Recovery of XylitolDocument5 pagesImproved Upstream Processing For Detoxification and Recovery of XylitolMARIA CAMILA MU�OZ COMETANo ratings yet
- Mesocarpo de Coco Verde Pretratado Por Un Proceso Alcalino Como Materia Prima para La Producción de BioetanolDocument39 pagesMesocarpo de Coco Verde Pretratado Por Un Proceso Alcalino Como Materia Prima para La Producción de BioetanolNancy EspinosaNo ratings yet
- Optimization of Chemical Pretreatments Using Response Surface Methodology For Second-Generation Ethanol Production From Coffee Husk WasteDocument13 pagesOptimization of Chemical Pretreatments Using Response Surface Methodology For Second-Generation Ethanol Production From Coffee Husk WasteAngie Estefania Rivera CordobaNo ratings yet
- Yang2015 PDFDocument11 pagesYang2015 PDFMuhammadFakhrizalFahmiNo ratings yet
- Faculty of Applied SciencesDocument39 pagesFaculty of Applied SciencesashNo ratings yet
- GSJ7016 (Aryan - Ramteke)Document20 pagesGSJ7016 (Aryan - Ramteke)CH24Aryan.RamtekeNo ratings yet
- Bietanol Kulit Pisang PDFDocument15 pagesBietanol Kulit Pisang PDFRomauli NataliaNo ratings yet
- Comparison of Volatile Components in Raw and Cooked Green Beans by GC-MS Using Dynamic Headspace Sampling and Microwave DesorptionDocument5 pagesComparison of Volatile Components in Raw and Cooked Green Beans by GC-MS Using Dynamic Headspace Sampling and Microwave DesorptionMahir YilmazNo ratings yet
- Extraction of Ethanol From CelluloseDocument20 pagesExtraction of Ethanol From CellulosePriyanka NagpureNo ratings yet
- Energies: Optimization of The Dilute Acid Hydrolyzator For Cellulose-to-Bioethanol SaccharificationDocument23 pagesEnergies: Optimization of The Dilute Acid Hydrolyzator For Cellulose-to-Bioethanol SaccharificationainmnrhNo ratings yet
- Study of The Hydrolysis of Sugar Cane Bagasse Using Phosphoric AcidDocument11 pagesStudy of The Hydrolysis of Sugar Cane Bagasse Using Phosphoric AcidRené MartínezNo ratings yet
- Isolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilDocument8 pagesIsolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilATSNo ratings yet
- Mepe14-Bio Fuels: Unit - IiiDocument22 pagesMepe14-Bio Fuels: Unit - IiiKrishna GummadiNo ratings yet
- Research Article: Enhanced Ethanol and Biogas Production From Pinewood by NMMO Pretreatment and Detailed Biomass AnalysisDocument11 pagesResearch Article: Enhanced Ethanol and Biogas Production From Pinewood by NMMO Pretreatment and Detailed Biomass AnalysislailaNo ratings yet
- Xilosa 1Document6 pagesXilosa 1Maria Victoria Ortega RamosNo ratings yet
- Bio-Based SolventsFrom EverandBio-Based SolventsFrançois JérômeNo ratings yet
- LeDoux Et Al 2016Document13 pagesLeDoux Et Al 2016Maria Victoria Ortega RamosNo ratings yet
- Jenny, Bingham, Padilla-Saravia. 1948. Nitrogen and Organic Matter Contents of Equatorial Soils of Colombia, South AmericaDocument14 pagesJenny, Bingham, Padilla-Saravia. 1948. Nitrogen and Organic Matter Contents of Equatorial Soils of Colombia, South AmericaMaria Victoria Ortega RamosNo ratings yet
- Bioconversion of Posthydrolysed Autohydrolysis Liquors: An Alternative For Xylitol Production From Corn CobsDocument8 pagesBioconversion of Posthydrolysed Autohydrolysis Liquors: An Alternative For Xylitol Production From Corn CobsMaria Victoria Ortega RamosNo ratings yet
- Jenny. 1948. Great Soil Groups in The Equatorial Regions of ColombiaDocument24 pagesJenny. 1948. Great Soil Groups in The Equatorial Regions of ColombiaMaria Victoria Ortega RamosNo ratings yet
- Sanchez. 2019. Properties and Management of Soils in The TropicsDocument686 pagesSanchez. 2019. Properties and Management of Soils in The TropicsMaria Victoria Ortega RamosNo ratings yet
- Xilosa 1Document6 pagesXilosa 1Maria Victoria Ortega RamosNo ratings yet
- Xilitol-Lecho FluidizadoDocument5 pagesXilitol-Lecho FluidizadoMaria Victoria Ortega RamosNo ratings yet
- 23Document11 pages23foad-7No ratings yet
- Fuel Processing Technology: Nalladurai Kaliyan, R. Vance MoreyDocument7 pagesFuel Processing Technology: Nalladurai Kaliyan, R. Vance MoreyMaria Victoria Ortega RamosNo ratings yet
- Fuel Processing Technology: Nalladurai Kaliyan, R. Vance MoreyDocument7 pagesFuel Processing Technology: Nalladurai Kaliyan, R. Vance MoreyMaria Victoria Ortega RamosNo ratings yet
- Microbial Xylitol Production From Corn Cobs Using Candida MagnoliaeDocument3 pagesMicrobial Xylitol Production From Corn Cobs Using Candida MagnoliaeMaria Victoria Ortega RamosNo ratings yet
- Charla 13. Agitación, Mezclado y Fluidos No NewtonianosDocument33 pagesCharla 13. Agitación, Mezclado y Fluidos No NewtonianosMaria Victoria Ortega RamosNo ratings yet
- Charla 13. Agitación, Mezclado y Fluidos No NewtonianosDocument33 pagesCharla 13. Agitación, Mezclado y Fluidos No NewtonianosMaria Victoria Ortega RamosNo ratings yet
- Charla 13. Agitación, Mezclado y Fluidos No NewtonianosDocument33 pagesCharla 13. Agitación, Mezclado y Fluidos No NewtonianosMaria Victoria Ortega RamosNo ratings yet
- A 09Document15 pagesA 09Mohamed MohsenNo ratings yet
- Akerholm - Characterization of The Crystalline Structure of Cellulose Using Static and Dynamic FT IR Spectroscopy - 2004Document10 pagesAkerholm - Characterization of The Crystalline Structure of Cellulose Using Static and Dynamic FT IR Spectroscopy - 2004Marcela TiboniNo ratings yet
- Schindler's List Theme Sheet Music For Piano, Violin (Solo)Document1 pageSchindler's List Theme Sheet Music For Piano, Violin (Solo)Sara SzaboNo ratings yet
- Income Tax Law & Practice Code: BBA-301 Unit - 2: Ms. Manisha Sharma Asst. ProfessorDocument32 pagesIncome Tax Law & Practice Code: BBA-301 Unit - 2: Ms. Manisha Sharma Asst. ProfessorVasu NarangNo ratings yet
- Medical Tourism A Look at How Medical Outsourcing Can Reshape Health Care - 2014Document18 pagesMedical Tourism A Look at How Medical Outsourcing Can Reshape Health Care - 2014avijitcu2007No ratings yet
- Artificial Intelligence and Applications: Anuj Gupta, Ankur BhadauriaDocument8 pagesArtificial Intelligence and Applications: Anuj Gupta, Ankur BhadauriaAnuj GuptaNo ratings yet
- Fisher C1 Series Pneumatic Controllers and TransmittersDocument12 pagesFisher C1 Series Pneumatic Controllers and TransmittersAnderson KundeNo ratings yet
- KUTAI MCCB Circuit Breaker Transfer Switch With Remote Monitoring OptionsDocument11 pagesKUTAI MCCB Circuit Breaker Transfer Switch With Remote Monitoring OptionsJanice LuNo ratings yet
- ADM2341 CH 8 Capstone QDocument2 pagesADM2341 CH 8 Capstone QjuiceNo ratings yet
- TIDCP vs. Manalang-DemigilioDocument2 pagesTIDCP vs. Manalang-DemigilioDi ko alam100% (1)
- MS For Surface Rock Bolts and Dowels (Rev-2)Document23 pagesMS For Surface Rock Bolts and Dowels (Rev-2)shahzadNo ratings yet
- Pravin Raut Sanjay Raut V EDDocument122 pagesPravin Raut Sanjay Raut V EDSandeep DashNo ratings yet
- Analog Circuits - IDocument127 pagesAnalog Circuits - IdeepakpeethambaranNo ratings yet
- BBA - Study On Gold As A Safer Investment CommodityDocument94 pagesBBA - Study On Gold As A Safer Investment CommoditySANJU GNo ratings yet
- In-N-Out vs. DoorDashDocument16 pagesIn-N-Out vs. DoorDashEaterNo ratings yet
- Crop Insurance Proposal 31.07.2023 (CLN) PCPMD Inputs Rev SCIDocument14 pagesCrop Insurance Proposal 31.07.2023 (CLN) PCPMD Inputs Rev SCIĐỗ ThăngNo ratings yet
- Valuation of Bonds and Stocks: Financial Management Prof. Deepa IyerDocument49 pagesValuation of Bonds and Stocks: Financial Management Prof. Deepa IyerAahaanaNo ratings yet
- PDFs/Sprinker NFPA 13/plan Review Sprinkler Checklist 13Document5 pagesPDFs/Sprinker NFPA 13/plan Review Sprinkler Checklist 13isbtanwir100% (1)
- Liebert Exs 10 20 Kva Brochure EnglishDocument8 pagesLiebert Exs 10 20 Kva Brochure Englishenrique espichan coteraNo ratings yet
- VDO Gauge - VL Hourmeter 12 - 24Document2 pagesVDO Gauge - VL Hourmeter 12 - 24HanNo ratings yet
- SEBI Circular On Online Processing of Investor Service Requests and Complaints by RTAs - June 8, 2023Document5 pagesSEBI Circular On Online Processing of Investor Service Requests and Complaints by RTAs - June 8, 2023Diya PandeNo ratings yet
- TechRef SoftstarterDocument11 pagesTechRef SoftstarterCesarNo ratings yet
- Pro & Contra Hydropower: AdvantagesDocument16 pagesPro & Contra Hydropower: AdvantagesbarukomkssNo ratings yet
- Hira Gaurd HouseDocument14 pagesHira Gaurd HouseNagadevan GovindanNo ratings yet
- EE 4702 Take-Home Pre-Final Questions: SolutionDocument11 pagesEE 4702 Take-Home Pre-Final Questions: SolutionmoienNo ratings yet
- ZF SI 6hp Family Tree enDocument2 pagesZF SI 6hp Family Tree enfercho573No ratings yet
- Wheel Loader Manual Agrison PDFDocument138 pagesWheel Loader Manual Agrison PDFTravisReign Dicang02No ratings yet
- 6063 Grade Aluminum ProfileDocument3 pages6063 Grade Aluminum ProfileVJ QatarNo ratings yet
- 01 04 2018Document55 pages01 04 2018sagarNo ratings yet
- Computer Systems Servicing NC II CGDocument238 pagesComputer Systems Servicing NC II CGRickyJeciel100% (2)
- Crime MappingDocument13 pagesCrime MappingRea Claire QuimnoNo ratings yet
- File System ImplementationDocument35 pagesFile System ImplementationSát Thủ Vô TìnhNo ratings yet