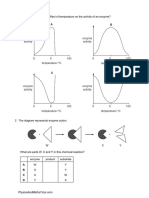
You might also like
- Pnas00043 0562Document5 pagesPnas00043 0562api-295783378No ratings yet
- Tetrahydrocannabinol and Endogenous Cannabinoid Anandamide Directly Potentiate The Function of Glycine ReceptorsDocument7 pagesTetrahydrocannabinol and Endogenous Cannabinoid Anandamide Directly Potentiate The Function of Glycine ReceptorsCarlos RodriguezNo ratings yet
- Energy Conservation by Succinate Decarboxylation in Veillonezla PandaDocument5 pagesEnergy Conservation by Succinate Decarboxylation in Veillonezla PandaMuhamad Alim MakhrusNo ratings yet
- Histamina h3Document7 pagesHistamina h3Nelly Solange TamayoNo ratings yet
- Update of KetamineDocument12 pagesUpdate of KetamineThoriqotil Haqqul MauludiyahNo ratings yet
- Navigation Search THC (Disambiguation) : TetrahydrocannabinolDocument14 pagesNavigation Search THC (Disambiguation) : TetrahydrocannabinolVictor Vacariu100% (1)
- Bioorganic & Medicinal Chemistry Letters: A A B C A D e B ADocument30 pagesBioorganic & Medicinal Chemistry Letters: A A B C A D e B ABi LouNo ratings yet
- 8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseDocument5 pages8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseSean Christopher PawlowskiNo ratings yet
- Tingenone NO Pathway Activation in Mice - EurJPharmacolDocument5 pagesTingenone NO Pathway Activation in Mice - EurJPharmacolRenata MendesNo ratings yet
- Formukong 1989Document5 pagesFormukong 1989ovidorlimagabrielNo ratings yet
- Clinical Pharmacokinetics of N-AcetylcysteineDocument12 pagesClinical Pharmacokinetics of N-AcetylcysteineTisaNo ratings yet
- Protective Role of Melatonin and Taurine Against Carbamazepine-Induced Toxicity in Freshly Isolated Rat HepatocytesDocument9 pagesProtective Role of Melatonin and Taurine Against Carbamazepine-Induced Toxicity in Freshly Isolated Rat HepatocytesOscar GuillenNo ratings yet
- Synthesis and Evaluation of Benzothiazole-Based Analogues As Novel, Potent, and Selective Fatty Acid Amide Hydrolase InhibitorsDocument11 pagesSynthesis and Evaluation of Benzothiazole-Based Analogues As Novel, Potent, and Selective Fatty Acid Amide Hydrolase InhibitorsVINODNo ratings yet
- High Affinity of The Naturally-Occurring Biflavonoid, Amentoflavon, To Brain Benzodiazepine Receptors in VitroDocument3 pagesHigh Affinity of The Naturally-Occurring Biflavonoid, Amentoflavon, To Brain Benzodiazepine Receptors in VitroJuan Carlos LrNo ratings yet
- Dermorphin Related PeptidesDocument5 pagesDermorphin Related PeptidesKambo VivaNo ratings yet
- J Bioorg 2020 103633Document13 pagesJ Bioorg 2020 103633Kumar NeerajNo ratings yet
- p-Nitrophenol-a-D-Glucopyranoside As Substrate For MeasurementDocument4 pagesp-Nitrophenol-a-D-Glucopyranoside As Substrate For MeasurementSarah Zielda NajibNo ratings yet
- Ijppr - Human 53 - 6 20 17 06 2020Document8 pagesIjppr - Human 53 - 6 20 17 06 2020Pharmacology CiuNo ratings yet
- Y-Aminobutyric: Comparison Analogues ONDocument7 pagesY-Aminobutyric: Comparison Analogues ONUmdatul MufiidahNo ratings yet
- Synthesis and Antimuscarinic Activity of 2-WEthyl-N-lhydroxyethylaminoethyl 22diphenylpropionate A Metaboliteof Aprophen jps.2600820603Document2 pagesSynthesis and Antimuscarinic Activity of 2-WEthyl-N-lhydroxyethylaminoethyl 22diphenylpropionate A Metaboliteof Aprophen jps.2600820603THEUSER0001No ratings yet
- C7ef PDFDocument3 pagesC7ef PDFVishak VsNo ratings yet
- J. S. Fowler Et Al - ( 11-C) Spiroperidol: Synthesis, Specific Activity Determination, and Biodistribution in MiceDocument9 pagesJ. S. Fowler Et Al - ( 11-C) Spiroperidol: Synthesis, Specific Activity Determination, and Biodistribution in MiceLonkesNo ratings yet
- Pnas00658 0499Document3 pagesPnas00658 0499abril ibarraNo ratings yet
- Cancer Prostate Cancer and Kalicreins 2 and 3 1997Document3 pagesCancer Prostate Cancer and Kalicreins 2 and 3 1997gustavopsoNo ratings yet
- ShraddhaDocument10 pagesShraddhashraddhapatil746No ratings yet
- 1414 431X BJMBR 1414 431X20176346Document8 pages1414 431X BJMBR 1414 431X20176346Guilherme Vasconcelos VasconcelosNo ratings yet
- Synthesis and Pain Inhibition Activity of The Analogs of 1-Allyl-3-Benzoylthiourea For New Analgesic Lead Compound DiscoveryDocument4 pagesSynthesis and Pain Inhibition Activity of The Analogs of 1-Allyl-3-Benzoylthiourea For New Analgesic Lead Compound Discoverymita ayu arditaNo ratings yet
- Enzimology GehrardDocument7 pagesEnzimology Gehrarddioneaguiar DioneNo ratings yet
- ColinergicosDocument12 pagesColinergicosELIAS YOSAFATH HERNANDEZ HERNANDEZNo ratings yet
- "Cannabinoids in The Treatment of Epilepsy" - New England Journal of MedicineDocument11 pages"Cannabinoids in The Treatment of Epilepsy" - New England Journal of MedicineloomcNo ratings yet
- Determination of Glyoxylyl-Peptide Concentration Using Oxime Chemistry and RP-HPLC AnalysisDocument5 pagesDetermination of Glyoxylyl-Peptide Concentration Using Oxime Chemistry and RP-HPLC AnalysisBenaoNo ratings yet
- 1980 Marden InvolvementDocument8 pages1980 Marden Involvementspanners 12No ratings yet
- Acetaminophen Prodrugs: Synthesis, Physicochemical Properties, and Analgesic ActivityDocument7 pagesAcetaminophen Prodrugs: Synthesis, Physicochemical Properties, and Analgesic ActivityNguyễn HàNo ratings yet
- The Efficiency of Glyceryl Behenate As Sustained-Release Agent Compared With Hydroxypropylcellulose in TabletsDocument7 pagesThe Efficiency of Glyceryl Behenate As Sustained-Release Agent Compared With Hydroxypropylcellulose in TabletsswabrijNo ratings yet
- Asymmetric Syntheses of Both Enantiomers of Amphetamine Hydrochloride Via Bakers' Yeast Reduction of PhenylacetoneDocument3 pagesAsymmetric Syntheses of Both Enantiomers of Amphetamine Hydrochloride Via Bakers' Yeast Reduction of PhenylacetoneLiNo ratings yet
- Maelicke2000 PDFDocument6 pagesMaelicke2000 PDFAllen SodaNo ratings yet
- Anesth Analg-2009-Habibi-Asl-936-42Document7 pagesAnesth Analg-2009-Habibi-Asl-936-42Mehboob AlamNo ratings yet
- Action of Antiepileptic and Anaesthetic Drugs On Na-And Ca-Spikes in Mammalian Non-Myelinated AxonsDocument9 pagesAction of Antiepileptic and Anaesthetic Drugs On Na-And Ca-Spikes in Mammalian Non-Myelinated AxonsSawita KumariNo ratings yet
- Evaluation of The Antioxidant and Prooxidant Actions of Gallic Acid and Its DerivativesDocument6 pagesEvaluation of The Antioxidant and Prooxidant Actions of Gallic Acid and Its DerivativesLourdes Pérez LópezNo ratings yet
- Luminescent ATP Detection Assay Kit Ab113849 Luminescent ATP Detection Assay Kit Ab113849Document4 pagesLuminescent ATP Detection Assay Kit Ab113849 Luminescent ATP Detection Assay Kit Ab113849carlosNo ratings yet
- Preclinical Evaluation of A New, Stabilized Neurotensin (8 - 13) Pseudopeptide Radiolabeled With TCDocument10 pagesPreclinical Evaluation of A New, Stabilized Neurotensin (8 - 13) Pseudopeptide Radiolabeled With TCtsengdNo ratings yet
- G. Keith Mulholland et al- Synthesis, Rodent Biodistribution, Dosimetry, Metabolism, and Monkey Images of Carbon-11- Labeled (+)-2alpha-Tropanyl Benzilate: A Central Muscarinic Receptor Imaging AgentDocument8 pagesG. Keith Mulholland et al- Synthesis, Rodent Biodistribution, Dosimetry, Metabolism, and Monkey Images of Carbon-11- Labeled (+)-2alpha-Tropanyl Benzilate: A Central Muscarinic Receptor Imaging AgentHumiopNo ratings yet
- Interaction of Acamprosate With Ethanol and Spermine On NMDA Receptors in Primary Cultured NeuronsDocument11 pagesInteraction of Acamprosate With Ethanol and Spermine On NMDA Receptors in Primary Cultured Neuronsapi-3851462No ratings yet
- Hit To LeadDocument4 pagesHit To LeadSatendra KumarNo ratings yet
- Erythrina Mulungu: Alkaloids Are Potent Inhibitors of Neuronal Nicotinic Receptor Currents in Mammalian CellsDocument6 pagesErythrina Mulungu: Alkaloids Are Potent Inhibitors of Neuronal Nicotinic Receptor Currents in Mammalian CellsgabrielledireitoNo ratings yet
- Ibarra-Lecue Et Al (2018) - Mecanismo Molecular de Uso Cannabis y EsquizofreniaDocument8 pagesIbarra-Lecue Et Al (2018) - Mecanismo Molecular de Uso Cannabis y EsquizofreniaDaniel LeinadNo ratings yet
- Marine Indole Alkaloids:Potencial New Drug Leads For The Control of Depression and AnxietyDocument26 pagesMarine Indole Alkaloids:Potencial New Drug Leads For The Control of Depression and AnxietyParth JadavNo ratings yet
- 1983 - Adrenergic Regulation of Adipocyte MetabolismDocument22 pages1983 - Adrenergic Regulation of Adipocyte MetabolismRaymond LaBoyNo ratings yet
- 10 1016@j Mex 2018 07 013Document10 pages10 1016@j Mex 2018 07 013Miy AichNo ratings yet
- Bonnot 1991Document2 pagesBonnot 1991weslyo98No ratings yet
- Proteinase K From Engyodontium Album: P-ChloromercuribenzoateDocument2 pagesProteinase K From Engyodontium Album: P-ChloromercuribenzoateviveksheteNo ratings yet
- Ketamine: Ketamine Is 2 - (O-Chlorophenyl) - 2 - (Methylamino) Cyclohexanone (Hydrochloride)Document15 pagesKetamine: Ketamine Is 2 - (O-Chlorophenyl) - 2 - (Methylamino) Cyclohexanone (Hydrochloride)Astrid Ghea GayatriNo ratings yet
- Sporo Genes'Document4 pagesSporo Genes'millodoriaNo ratings yet
- Tingenone, A Pentacyclic Triterpene, Induces Peripheral Antinociception Due To Cannabinoid Receptors Activation in Mice - InflammopharmacologyDocument9 pagesTingenone, A Pentacyclic Triterpene, Induces Peripheral Antinociception Due To Cannabinoid Receptors Activation in Mice - InflammopharmacologyRenata MendesNo ratings yet
- tmp6666 TMPDocument9 pagestmp6666 TMPFrontiersNo ratings yet
- CapsaicinDocument12 pagesCapsaicinExlonk Gil PeláezNo ratings yet
- Benzimidazolyl ChalconeDocument9 pagesBenzimidazolyl Chalconeg20kpNo ratings yet
- Malterud 2000Document5 pagesMalterud 2000redhalfi.fadhilaNo ratings yet
- CholinesteraseDocument2 pagesCholinesteraseGeorge-Alexandru MarinescuNo ratings yet
- A Study of The Larvicidal Activity of Two Croton Species From Northeastern Brazil Against Aedes AegyptiDocument6 pagesA Study of The Larvicidal Activity of Two Croton Species From Northeastern Brazil Against Aedes AegyptiJohn MaddenNo ratings yet
- NIH Public Access: Author ManuscriptDocument16 pagesNIH Public Access: Author ManuscriptJohn MaddenNo ratings yet
- Zitek Et Al Oct 2020Document17 pagesZitek Et Al Oct 2020John MaddenNo ratings yet
- Drug and Alcohol DependenceDocument8 pagesDrug and Alcohol DependenceJohn MaddenNo ratings yet
- Anis Et Al Jan 2021Document14 pagesAnis Et Al Jan 2021John MaddenNo ratings yet
- Pharmacology, Biochemistry and BehaviorDocument9 pagesPharmacology, Biochemistry and BehaviorJohn MaddenNo ratings yet
- Antibacterial Cannabinoids From Cannabis Sativa - Appendino - J. Nat. Prod 71 (2008)Document4 pagesAntibacterial Cannabinoids From Cannabis Sativa - Appendino - J. Nat. Prod 71 (2008)dextroenantiomerNo ratings yet
- Kapeghian Et Al 1983Document3 pagesKapeghian Et Al 1983John MaddenNo ratings yet
- Izzo Et Al Jun 2012Document17 pagesIzzo Et Al Jun 2012John MaddenNo ratings yet
- Ligresti Et Al Sep 2006Document13 pagesLigresti Et Al Sep 2006John MaddenNo ratings yet
- Ilan Et Al Sep 2005Document11 pagesIlan Et Al Sep 2005John MaddenNo ratings yet
- Dell W2306C LCD Monitor Service ManualDocument104 pagesDell W2306C LCD Monitor Service ManualIsrael B ChavezNo ratings yet
- Ask A Monk EnlightenmentDocument16 pagesAsk A Monk EnlightenmentPetruoka EdmundasNo ratings yet
- 04 SAMSS 005 Check ValvesDocument9 pages04 SAMSS 005 Check ValvesShino UlahannanNo ratings yet
- All Pop SongsDocument53 pagesAll Pop SongsMadeleneQuiogueNo ratings yet
- Comparative Performance of Some Cattle Breeds Under Barani Conditions of PakistanDocument4 pagesComparative Performance of Some Cattle Breeds Under Barani Conditions of PakistanMasood HassanNo ratings yet
- Upaam 1135891 202105060749199700Document18 pagesUpaam 1135891 202105060749199700Kartik KapoorNo ratings yet
- Genie GS-1930 Parts ManualDocument194 pagesGenie GS-1930 Parts ManualNestor Matos GarcíaNo ratings yet
- Paranoia XP - Gamemaster Screen Booklet - Mandatory Fun Enforcement PackDocument24 pagesParanoia XP - Gamemaster Screen Booklet - Mandatory Fun Enforcement PackStBash100% (3)
- Time Value of Money LectureDocument54 pagesTime Value of Money LectureRanin, Manilac Melissa SNo ratings yet
- Multiple Choice Enzymes Plant and Animal NutritionDocument44 pagesMultiple Choice Enzymes Plant and Animal Nutritionliufanjing07No ratings yet
- Greater Occipital Nerve Block: Pain ManagementDocument3 pagesGreater Occipital Nerve Block: Pain Managementkillingeyes177No ratings yet
- Sheet Metal Manufacturing Companies NoidaDocument9 pagesSheet Metal Manufacturing Companies NoidaAmanda HoldenNo ratings yet
- Superposition and Statically Indetermina - GDLCDocument25 pagesSuperposition and Statically Indetermina - GDLCAnonymous frFFmeNo ratings yet
- Intel Stratix 10 Avalon - MM Interface For PCI Express Solutions User GuideDocument173 pagesIntel Stratix 10 Avalon - MM Interface For PCI Express Solutions User Guideenoch richardNo ratings yet
- Chemistry Notes: SUBJECT: Leaving Cert Chemistry Level: TEACHER: Tara LyonsDocument5 pagesChemistry Notes: SUBJECT: Leaving Cert Chemistry Level: TEACHER: Tara LyonsSevinc NuriyevaNo ratings yet
- Ge Fairchild Brochure PDFDocument2 pagesGe Fairchild Brochure PDFDharmesh patelNo ratings yet
- Chapter 10 - The Mature ErythrocyteDocument55 pagesChapter 10 - The Mature ErythrocyteSultan AlexandruNo ratings yet
- Siprotec 7ut82 ProfileDocument2 pagesSiprotec 7ut82 ProfileOliver Atahuichi TorrezNo ratings yet
- Contact Inform 2002 PDFDocument24 pagesContact Inform 2002 PDFFrank AlmeidaNo ratings yet
- QuantAssay Software Manual 11-Mar-2019Document51 pagesQuantAssay Software Manual 11-Mar-2019LykasNo ratings yet
- Hot Topic 02 Good Light Magazine 56smDocument24 pagesHot Topic 02 Good Light Magazine 56smForos IscNo ratings yet
- Cac de Luyen ThiDocument18 pagesCac de Luyen ThiVinh Dao CongNo ratings yet
- Eating With Chloe Lets EatDocument150 pagesEating With Chloe Lets Eatemily.jarrodNo ratings yet
- The Wilson Quarterly: The World's New Numbers by Martin WalkerDocument13 pagesThe Wilson Quarterly: The World's New Numbers by Martin WalkerDavid WeekNo ratings yet
- VTA28-G5: Fuel OptimizedDocument3 pagesVTA28-G5: Fuel OptimizedIslam HemdanNo ratings yet
- THE WEATHER LISTENING - Stalin GuañunaDocument1 pageTHE WEATHER LISTENING - Stalin GuañunaSTALIN JESUS GUA�UNA CHICAIZANo ratings yet
- Aesculap Saw GD307 - Service ManualDocument16 pagesAesculap Saw GD307 - Service ManualFredi PançiNo ratings yet
- Germania Agenturinfo enDocument4 pagesGermania Agenturinfo enДушан СтепановићNo ratings yet
- In-Service Welding of Pipelines Industry Action PlanDocument13 pagesIn-Service Welding of Pipelines Industry Action Planعزت عبد المنعم100% (1)
- Firestone & Scholl - Cognition Does Not Affect Perception, Evaluating Evidence For Top-Down EffectsDocument77 pagesFirestone & Scholl - Cognition Does Not Affect Perception, Evaluating Evidence For Top-Down EffectsRed JohnNo ratings yet