You might also like
- New Oxford General Knowledge TG 1Document70 pagesNew Oxford General Knowledge TG 1abia sajidNo ratings yet
- Donna Haraway - Manifiesto CyborgDocument36 pagesDonna Haraway - Manifiesto CyborgAyelen BirrielNo ratings yet
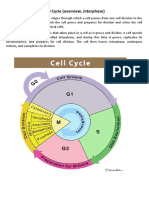
- Ii M.Sc. Botany Cell and Molecular Biology Unit: Ii Cell CycleDocument36 pagesIi M.Sc. Botany Cell and Molecular Biology Unit: Ii Cell CycleDeblina JanaNo ratings yet
- Unit 1Document52 pagesUnit 1Dsce BtNo ratings yet
- Cell CycleDocument4 pagesCell CycleSujith KuttanNo ratings yet
- Cell Cycle: (Fundamental Process)Document8 pagesCell Cycle: (Fundamental Process)r_borresNo ratings yet
- Cell Cycle and CheckpointsDocument12 pagesCell Cycle and Checkpointsjayyyyyrg16No ratings yet
- Cell Cycle & DivisionDocument28 pagesCell Cycle & DivisionThelma SimonNo ratings yet
- Cell CycleDocument15 pagesCell CycleFaheem ullahNo ratings yet
- Cell ModificationsDocument8 pagesCell ModificationsKriezhel Kim TadleNo ratings yet
- Cell Division and Genetic Inheritance - FormattedDocument24 pagesCell Division and Genetic Inheritance - FormattedAISHA MUHAMMADNo ratings yet
- Cell Cycle (Scitable)Document2 pagesCell Cycle (Scitable)LibrofiloNo ratings yet
- Cell CycleDocument10 pagesCell CycleDrexel DalaygonNo ratings yet
- 2 Cell CycleDocument31 pages2 Cell Cyclemuharramisalma009No ratings yet
- Cell DivisionDocument4 pagesCell DivisionHajie RosarioNo ratings yet
- Cell CycleDocument9 pagesCell CycleTejaswini AvusulaNo ratings yet
- Cell Cycle: - Dr. Ishita SinghalDocument69 pagesCell Cycle: - Dr. Ishita SinghalDR. ISHITA SINGHAL100% (1)
- Cell Cycle, Mitosis, Chaper12Document24 pagesCell Cycle, Mitosis, Chaper12prehealthhelp43% (7)
- Cell CycleDocument19 pagesCell CycleAnahita SuriNo ratings yet
- Bio 1Document13 pagesBio 1Mary Ann FriasNo ratings yet
- Lesson 6 - Cell Cycle and CheckpointsDocument6 pagesLesson 6 - Cell Cycle and CheckpointsReuben Al'jaldi100% (1)
- Cell Cycle CheckpointsDocument6 pagesCell Cycle CheckpointsJoyce NoblezaNo ratings yet
- An Overview of Cell Cycle (Lec # 3)Document6 pagesAn Overview of Cell Cycle (Lec # 3)Izhar HussainNo ratings yet
- Cell Cycle ReviewDocument21 pagesCell Cycle Reviewdaniela Hernandez LopezNo ratings yet
- Cell CycleDocument9 pagesCell Cyclestevensb055No ratings yet
- M2L1 ActivateDocument2 pagesM2L1 ActivateVenice De los ReyesNo ratings yet
- Cell Cycle and Cell DivisionDocument39 pagesCell Cycle and Cell DivisionSayantan SethNo ratings yet
- Lesson 6 Cell Cycle HAND OUTDocument10 pagesLesson 6 Cell Cycle HAND OUTAyera DicaliNo ratings yet
- Cell CycleDocument15 pagesCell CycleRhea GulayNo ratings yet
- CBSE-XI Biology - Chap-10-12Document32 pagesCBSE-XI Biology - Chap-10-12krishnabhavith3No ratings yet
- Cell DivisionDocument20 pagesCell DivisionABHINABA GUPTANo ratings yet
- Gen Biology UNIT 2 - Lesson 1Document8 pagesGen Biology UNIT 2 - Lesson 1Albert Jade Pontimayor LegariaNo ratings yet
- Cellcycle-190703053908Document68 pagesCellcycle-190703053908AyeshaNo ratings yet
- Cell CycleDocument22 pagesCell CycleMUHAMMAD KHANNo ratings yet
- Cell Bio AssignmentDocument11 pagesCell Bio AssignmentPranay PatnaikNo ratings yet
- Cell Cycle and DivisionDocument47 pagesCell Cycle and DivisionMadhura ShekatkarNo ratings yet
- Cell BiologyDocument24 pagesCell BiologySheelendra Mangal BhattNo ratings yet
- Handouts Gen. Bio Oct 3 6 2022Document14 pagesHandouts Gen. Bio Oct 3 6 2022Mark Kenneth CastilloNo ratings yet
- Lecture 1Document48 pagesLecture 1NIMRAHNo ratings yet
- Eukaryotic Cell CycleDocument96 pagesEukaryotic Cell CycleRaghav Oberoi100% (1)
- Cell CycleDocument2 pagesCell CycleJudith MabaoNo ratings yet
- The Cell CycleDocument4 pagesThe Cell CycleExcellent KhozaNo ratings yet
- Module 1Document48 pagesModule 1Saalif RahmanNo ratings yet
- Module 3 - Gen Bio 1 - MidtermDocument7 pagesModule 3 - Gen Bio 1 - MidtermAngel Cuacko GacmatanNo ratings yet
- Lab11 The Cell Cycle PDFDocument32 pagesLab11 The Cell Cycle PDFعبدالرحمن عدي عبدالفتاحNo ratings yet
- Cell Cycle Handouts - PDF (Highlighted)Document4 pagesCell Cycle Handouts - PDF (Highlighted)Shien VarronNo ratings yet
- Cell DivisionDocument11 pagesCell DivisionlaktshyamanigandanNo ratings yet
- Bio 1S The Cell CycleDocument92 pagesBio 1S The Cell Cyclestem.patinosamcharrieNo ratings yet
- UNIT 4 Cell CycleDocument24 pagesUNIT 4 Cell CycleChennille Ann Bleu GundayaoNo ratings yet
- IJSO Biology Module - 1Document137 pagesIJSO Biology Module - 1Ikhbaat Atiqur RehmanNo ratings yet
- Cell CycleDocument6 pagesCell CycleIgnacio, Moira Jomille K.No ratings yet
- 6 MitosisDocument4 pages6 Mitosisdevanshianand555No ratings yet
- Cell DivisionDocument40 pagesCell DivisionMohamed NourNo ratings yet
- Week 3 Modules Grade 12Document13 pagesWeek 3 Modules Grade 12Joe NasalitaNo ratings yet
- Module in EnglishDocument28 pagesModule in EnglishIannNo ratings yet
- Cell Cycle Regulation - Control Points (Part I)Document24 pagesCell Cycle Regulation - Control Points (Part I)Amara TargaryenNo ratings yet
- Cell CycleDocument31 pagesCell CycleMariah Missy DatuNo ratings yet
- Botany NCERT Cell CycleDocument11 pagesBotany NCERT Cell CycleVishal SorteyNo ratings yet
- Cell and Molecular BiologyDocument9 pagesCell and Molecular BiologyjnsenguptaNo ratings yet
- Cell DivisionDocument20 pagesCell DivisionSoumodip ChakrabortyNo ratings yet
- Hormones: Roshan Ali Assistant Professor Ibms, KmuDocument6 pagesHormones: Roshan Ali Assistant Professor Ibms, KmuUrdu KahaniNo ratings yet
- Cholera, Diphtheria, Pertusis, and Tetanus: Muhammad Iqbal Lecturer KMUDocument11 pagesCholera, Diphtheria, Pertusis, and Tetanus: Muhammad Iqbal Lecturer KMUUrdu KahaniNo ratings yet
- CommincationDocument31 pagesCommincationUrdu KahaniNo ratings yet
- Cholera, Diphtheria, Pertusis, and Tetanus: Muhammad Iqbal Lecturer KMUDocument11 pagesCholera, Diphtheria, Pertusis, and Tetanus: Muhammad Iqbal Lecturer KMUUrdu KahaniNo ratings yet
- 6nucleic Acids (INS)Document24 pages6nucleic Acids (INS)Urdu KahaniNo ratings yet
- Introduction To MicrobiologyDocument11 pagesIntroduction To MicrobiologyUrdu KahaniNo ratings yet
- Metabolism of Fats: Roshan Ali Assistant Professor Ibms, KmuDocument6 pagesMetabolism of Fats: Roshan Ali Assistant Professor Ibms, KmuUrdu KahaniNo ratings yet
- Advance Exercises On AdjDocument13 pagesAdvance Exercises On AdjUrdu KahaniNo ratings yet
- Unit - VIII (Concept of Teaching and Learning)Document39 pagesUnit - VIII (Concept of Teaching and Learning)Urdu Kahani100% (1)
- 11metabolism of Carbohydrates 2Document8 pages11metabolism of Carbohydrates 2Urdu KahaniNo ratings yet
- 9, Concept of Isolation Unit-VDocument9 pages9, Concept of Isolation Unit-VUrdu Kahani100% (1)
- Metabolism of Proteins: Roshan Ali Assistant Professor Ibms, KmuDocument46 pagesMetabolism of Proteins: Roshan Ali Assistant Professor Ibms, KmuUrdu KahaniNo ratings yet
- 6nucleic Acids (INS)Document27 pages6nucleic Acids (INS)Urdu KahaniNo ratings yet
- Blood: By: Mah-U-Din ROLL NO:10850 Degree: B.S Programme Deptt: MLT City University of Engineering and TechnologyDocument8 pagesBlood: By: Mah-U-Din ROLL NO:10850 Degree: B.S Programme Deptt: MLT City University of Engineering and TechnologyUrdu KahaniNo ratings yet
- Virus Properties Unit-I-1Document15 pagesVirus Properties Unit-I-1Urdu KahaniNo ratings yet
- Normal Flora unit-IVDocument23 pagesNormal Flora unit-IVUrdu KahaniNo ratings yet
- 5joints (Unit-V)Document22 pages5joints (Unit-V)Urdu KahaniNo ratings yet
- 2homeostasis and Adaptation (unit-II)Document20 pages2homeostasis and Adaptation (unit-II)Urdu KahaniNo ratings yet
- Armstrong 1991Document7 pagesArmstrong 1991annesNo ratings yet
- Jntu Kakinada - B.tech - Earthquake Resistant Design r13 Rt42013c042017 FR 130Document2 pagesJntu Kakinada - B.tech - Earthquake Resistant Design r13 Rt42013c042017 FR 130Vinod KumarNo ratings yet
- Literature Review CrosswordDocument4 pagesLiterature Review Crosswordaflsjcibl100% (1)
- Project Proposal CWTSDPMDocument6 pagesProject Proposal CWTSDPMJacel Reinard ArceoNo ratings yet
- West Philippines Institute of Tourism, Science, and TechnologyDocument13 pagesWest Philippines Institute of Tourism, Science, and TechnologyFrenchesca OrtinezNo ratings yet
- ITP - CCTV System InstallationDocument6 pagesITP - CCTV System InstallationGultom Juliana LiliesNo ratings yet
- Review of Related Literature of Potato BatteryDocument7 pagesReview of Related Literature of Potato Batteryafmzrxbdfadmdq100% (2)
- Week 1-Laq's ObDocument4 pagesWeek 1-Laq's ObAswin RajNo ratings yet
- A5e35857004-Agen Multi Oi En-UsDocument350 pagesA5e35857004-Agen Multi Oi En-UsAmir KalčoNo ratings yet
- Tuesday Geography PowerpointDocument13 pagesTuesday Geography PowerpointPetra BalázsyNo ratings yet
- Energy Science SyllabusDocument1 pageEnergy Science SyllabusAbhinav DwivediNo ratings yet
- DNB Vol29 NoSuppl 1 64Document9 pagesDNB Vol29 NoSuppl 1 64Shamsul ArefinNo ratings yet
- Seed Germination Final Lab ReportDocument6 pagesSeed Germination Final Lab Reportapi-509411193No ratings yet
- Jawaharlal Nehru Technological University Hyderabad: Date& Day R17 R16 R15 R13 R09Document1 pageJawaharlal Nehru Technological University Hyderabad: Date& Day R17 R16 R15 R13 R09TempNo ratings yet
- Drd-E Installation Instructions 01Document12 pagesDrd-E Installation Instructions 01Ha ssanNo ratings yet
- Ejercicios Probl Valor Frontera Barber8Document2 pagesEjercicios Probl Valor Frontera Barber8Exequiel SeguelNo ratings yet
- EnergyNext Vol 05 Issue 12 Oct 2015Document84 pagesEnergyNext Vol 05 Issue 12 Oct 2015IndiaNextNo ratings yet
- Coulomb's Law MC Questions PDFDocument7 pagesCoulomb's Law MC Questions PDFLynn Hollenbeck BreindelNo ratings yet
- ABRJ2014 VolumeIDocument154 pagesABRJ2014 VolumeIAmna AliNo ratings yet
- Aw Pair of Linear Equations Grade 10 (24-25)Document13 pagesAw Pair of Linear Equations Grade 10 (24-25)sana022009No ratings yet
- Brain-Mind Bulletin June-July 1994 Volume 19, Numbers 9-10Document8 pagesBrain-Mind Bulletin June-July 1994 Volume 19, Numbers 9-10paulcaubetNo ratings yet
- 1 - Ions WorksheetDocument2 pages1 - Ions Worksheetmuieed siddiqui100% (1)
- Astm B805Document7 pagesAstm B805Jonicus-DextoreNo ratings yet
- Introduction To Gender StudiesDocument5 pagesIntroduction To Gender StudiesAQSA JAMEELNo ratings yet
- DDocument4 pagesDUrfia RizqiNo ratings yet
- GeomphaseDocument83 pagesGeomphaseatticus peinNo ratings yet
- The Four of Effective: Pillars NegotiationDocument7 pagesThe Four of Effective: Pillars NegotiationMariusNo ratings yet
- Effects of Restricted Feed Intake On Heat Energy BDocument14 pagesEffects of Restricted Feed Intake On Heat Energy BdrcdevaNo ratings yet