You might also like
- Trypanosoma cruzi, the Causative Agent of Chagas' Disease: A Biological, Cultural, and Economic ReviewFrom EverandTrypanosoma cruzi, the Causative Agent of Chagas' Disease: A Biological, Cultural, and Economic ReviewNo ratings yet
- Origins of Syphilis and Management in The Immunocompetent Patient: Facts and ControversiesDocument6 pagesOrigins of Syphilis and Management in The Immunocompetent Patient: Facts and ControversiesegerpratamaNo ratings yet
- People, Parasites, and Plowshares: Learning From Our Body's Most Terrifying InvadersFrom EverandPeople, Parasites, and Plowshares: Learning From Our Body's Most Terrifying InvadersRating: 4.5 out of 5 stars4.5/5 (2)
- Treponema Pallidum: Download This Page in PDF Format (Can Be Old) TaxonomyDocument11 pagesTreponema Pallidum: Download This Page in PDF Format (Can Be Old) TaxonomyGregorio De Las CasasNo ratings yet
- American Trypanosomiasis Chagas Disease: One Hundred Years of ResearchFrom EverandAmerican Trypanosomiasis Chagas Disease: One Hundred Years of ResearchJenny TelleriaNo ratings yet
- 6.paleogenesis and Paleo-Epidemiology of Primate MalariaDocument25 pages6.paleogenesis and Paleo-Epidemiology of Primate Malarialaura isabellaNo ratings yet
- Black Fungi: Clinical and Pathogenic ApproachesDocument8 pagesBlack Fungi: Clinical and Pathogenic ApproachesমিনহাজসাকিবNo ratings yet
- The Paleoepidemiology of American....Document2 pagesThe Paleoepidemiology of American....laura isabellaNo ratings yet
- On The Age of LeprosyDocument8 pagesOn The Age of LeprosyNoraNo ratings yet
- This Week's Citation Classic® - : Science 193:673-5Document1 pageThis Week's Citation Classic® - : Science 193:673-5Budi LamakaNo ratings yet
- 2013 Yaws Seminar LancetDocument11 pages2013 Yaws Seminar LancetOlivia Halim KumalaNo ratings yet
- CDC 72296 DS1Document8 pagesCDC 72296 DS1Sam BothMongkulNo ratings yet
- Out 8Document19 pagesOut 8ZakiaaNo ratings yet
- 32 Arthropod Borne DiseasesDocument16 pages32 Arthropod Borne Diseasesvieira.barrosNo ratings yet
- Yellow Fever Term PaperDocument7 pagesYellow Fever Term Paperafdtuwxrb100% (1)
- Harbeck Et Al 2013 Yersinia Pestis DNA From Skeletal Remains From The 6th Century AD Reveals Insights Into Justinianic PlagueDocument8 pagesHarbeck Et Al 2013 Yersinia Pestis DNA From Skeletal Remains From The 6th Century AD Reveals Insights Into Justinianic PlagueTeresa rivasNo ratings yet
- Thesis Paper On SyphilisDocument7 pagesThesis Paper On Syphiliskulilev0bod3100% (3)
- Et Al. - On The Origin of LeprosyDocument15 pagesEt Al. - On The Origin of LeprosyRoberto SanchezNo ratings yet
- Systemic Mycoses in Dogs and CatsDocument9 pagesSystemic Mycoses in Dogs and CatsMarcoahrNo ratings yet
- Germs, Malaria and The Invention of Mansonian Tropical Medicine: From 'Diseases in The Tropics' To 'Tropical Diseases'Document27 pagesGerms, Malaria and The Invention of Mansonian Tropical Medicine: From 'Diseases in The Tropics' To 'Tropical Diseases'JOSE IGNACIO MOGROVEJO PALOMONo ratings yet
- Pathobiology of Cell Wall and Cell Signal Transduction: Pneumocystis Pneumonia: Life CycleDocument12 pagesPathobiology of Cell Wall and Cell Signal Transduction: Pneumocystis Pneumonia: Life CycleVanessa YanzaguanoNo ratings yet
- Basic Clinical Parasitology PDF 4656784Document1 pageBasic Clinical Parasitology PDF 4656784Natalie EnriquezNo ratings yet
- Epidemiology of Helicobacter Pylori InfectionDocument5 pagesEpidemiology of Helicobacter Pylori InfectionCarolina VillagarayNo ratings yet
- Ancient Pathogen GenomicsDocument2 pagesAncient Pathogen Genomicsyvonne syNo ratings yet
- Helminth Life Cycles: Dept. of Microbiology, SUNY Upstate Medical Center, SyracuseDocument10 pagesHelminth Life Cycles: Dept. of Microbiology, SUNY Upstate Medical Center, Syracusebelelmusa86.mhmbNo ratings yet
- The Plague of Athens: Epidemiology and PaleopathologyDocument13 pagesThe Plague of Athens: Epidemiology and PaleopathologyCherry HtunNo ratings yet
- Neurosifilis Berger2014Document12 pagesNeurosifilis Berger2014eva yustianaNo ratings yet
- Feral Pigeons As Carriers of Cryptococcus Laurentii, C. Uniguttulatus and Debaryomyces HanseniiDocument3 pagesFeral Pigeons As Carriers of Cryptococcus Laurentii, C. Uniguttulatus and Debaryomyces HanseniiTANIA CORONADO ROMERONo ratings yet
- Daedly DeasesrabiesDocument144 pagesDaedly Deasesrabieslepoete40No ratings yet
- Ilns 47 54-1Document22 pagesIlns 47 54-1João Filipe NducaNo ratings yet
- Development Team: Zoology Biology of Parasitism Morphology, Life Cycle, Mode of Infection of TrypanosomaDocument17 pagesDevelopment Team: Zoology Biology of Parasitism Morphology, Life Cycle, Mode of Infection of TrypanosomaIrfan Pathan KakarNo ratings yet
- Treponema Pallidum Pallidum PertenueDocument13 pagesTreponema Pallidum Pallidum PertenueCelta Gómez TrejoNo ratings yet
- The Johns Hopkins University Press, Classical Association of The Atlantic States The Classical WorldDocument6 pagesThe Johns Hopkins University Press, Classical Association of The Atlantic States The Classical Worldahmet mörelNo ratings yet
- Pneumocystis Carinii Pneumonia: A ReviewDocument10 pagesPneumocystis Carinii Pneumonia: A ReviewDiana GhawaNo ratings yet
- On Anatomical Disease : Canadian MedicalDocument9 pagesOn Anatomical Disease : Canadian Medicalvlad-bNo ratings yet
- Pnas 1317723111Document14 pagesPnas 1317723111IrinaNo ratings yet
- baker2015Document9 pagesbaker2015Wojciech WawrętyNo ratings yet
- Malaria EssayDocument4 pagesMalaria EssayKiril ManolovNo ratings yet
- What Is MalariaDocument6 pagesWhat Is MalariaequityfoundationNo ratings yet
- Evolution of Immune Genes Is Associated With The Black DeathDocument22 pagesEvolution of Immune Genes Is Associated With The Black DeathEduardo Pepe Melgarejo DíazNo ratings yet
- (Advances in Veterinary Science and Comparative Medicine 34) Richard A. McFeely (Eds.) - Domestic Animal Cytogenetics - Advances in Veterinary Science and Comparative Medicine (1990, Academic Press)Document317 pages(Advances in Veterinary Science and Comparative Medicine 34) Richard A. McFeely (Eds.) - Domestic Animal Cytogenetics - Advances in Veterinary Science and Comparative Medicine (1990, Academic Press)Jose Franco NaviaNo ratings yet
- Nihms 940015Document50 pagesNihms 940015Maria Salome Olortegui AceroNo ratings yet
- Parasitology: Tissue NemathodesDocument41 pagesParasitology: Tissue NemathodesSwar KlesNo ratings yet
- An. Bras. Dermatol. Vol.81 No.2 Rio de Janeiro Mar./Apr. 2006Document21 pagesAn. Bras. Dermatol. Vol.81 No.2 Rio de Janeiro Mar./Apr. 2006farah bachmidNo ratings yet
- Histoplasmosis in Dogs and Cats: Catharina Brömel, DR - Med.Vet., and Jane E. Sykes, BVSC (Hons), PHD, DacvimDocument6 pagesHistoplasmosis in Dogs and Cats: Catharina Brömel, DR - Med.Vet., and Jane E. Sykes, BVSC (Hons), PHD, DacvimdpcamposhNo ratings yet
- Seasonal Variation of Tungiasis in An Endemic CommDocument6 pagesSeasonal Variation of Tungiasis in An Endemic CommGiovanna RotondoNo ratings yet
- Huxsoll 1976Document12 pagesHuxsoll 1976Paula Andrea Castro MartinezNo ratings yet
- Gaston Naessens Somatid Life Cycle 1121Document4 pagesGaston Naessens Somatid Life Cycle 1121Cheshire CatNo ratings yet
- Rodgers 2015Document5 pagesRodgers 2015No LimitNo ratings yet
- 3 Protozoa1 RMDocument7 pages3 Protozoa1 RMRaihan Zulfiqar HalimNo ratings yet
- Plague: from natural disease to bioterrorismDocument9 pagesPlague: from natural disease to bioterrorismJuan Carlos GutiérrezNo ratings yet
- Surveillance of Potential Hosts and Vectors of Scrub Typhus in TaiwanDocument11 pagesSurveillance of Potential Hosts and Vectors of Scrub Typhus in TaiwanNhe FirmansyahNo ratings yet
- Medical Protozoology. 1Document55 pagesMedical Protozoology. 1miniwhiteyNo ratings yet
- Spaganosis Case ReviewDocument9 pagesSpaganosis Case ReviewRapatra ChanalertpatraNo ratings yet
- TrypanosomiasisDocument12 pagesTrypanosomiasisangel pindaNo ratings yet
- David A Warrell: Centre For Tropical Medicine, University of Oxford, Oxford, UKDocument3 pagesDavid A Warrell: Centre For Tropical Medicine, University of Oxford, Oxford, UKDelenda ChamchoumaNo ratings yet
- Comparative Immunology, Microbiology and Infectious DiseasesDocument7 pagesComparative Immunology, Microbiology and Infectious DiseasesDIEGO FERNANDO TULCAN SILVANo ratings yet
- The History, Usage, and Misinformation of Vaccines 1Document20 pagesThe History, Usage, and Misinformation of Vaccines 1api-534716659No ratings yet
- About Pleomorphism. Suppressed Medical KnowledgeDocument17 pagesAbout Pleomorphism. Suppressed Medical KnowledgeCarl MacCord50% (2)
- Vasilakis Et Al., 2019Document38 pagesVasilakis Et Al., 2019Lúcia ReisNo ratings yet
- Pare's Famous Medical SayingDocument3 pagesPare's Famous Medical Sayingorlandom61No ratings yet
- The Most Difficult Career - Women's History in ScienceDocument9 pagesThe Most Difficult Career - Women's History in Scienceorlandom61No ratings yet
- The History and Philosophy of Women in Science - A Review EssayDocument29 pagesThe History and Philosophy of Women in Science - A Review Essayorlandom61No ratings yet
- Rita - Profiles of Rita Levi-Montalcini, Ricardo Miledi, Simon LeVay, Erich Jarvis, and Steve RamirezDocument8 pagesRita - Profiles of Rita Levi-Montalcini, Ricardo Miledi, Simon LeVay, Erich Jarvis, and Steve Ramirezorlandom61No ratings yet
- Ambroise Paré - The Oculist's Eye - Connections Between Cataract Couching, Anatomy, and Visual Theory in The RenaissanceDocument16 pagesAmbroise Paré - The Oculist's Eye - Connections Between Cataract Couching, Anatomy, and Visual Theory in The Renaissanceorlandom61No ratings yet
- The History of Prosthetics from Ancient Egypt to Ambroise ParéDocument6 pagesThe History of Prosthetics from Ancient Egypt to Ambroise Paréorlandom61No ratings yet
- Ambroise Paré - IDocument1 pageAmbroise Paré - Iorlandom61No ratings yet
- Ambroise Paré - III - HernigouDocument6 pagesAmbroise Paré - III - Hernigouorlandom61No ratings yet
- Ambroise Paré - Premier Chirurgien Du Rol - The Life of Ambroise Pare (1510-1590)Document3 pagesAmbroise Paré - Premier Chirurgien Du Rol - The Life of Ambroise Pare (1510-1590)orlandom61No ratings yet
- Ambroise Paré - MDocument4 pagesAmbroise Paré - Morlandom61No ratings yet
- Ambroise Paré - Historical-Gleanings-1962Document3 pagesAmbroise Paré - Historical-Gleanings-1962orlandom61No ratings yet
- Ambroise Paré - ODocument3 pagesAmbroise Paré - Oorlandom61No ratings yet
- Ambroise Paré - eDocument3 pagesAmbroise Paré - eorlandom61No ratings yet
- Ambroise Paré - I - HernigouDocument5 pagesAmbroise Paré - I - Hernigouorlandom61No ratings yet
- Ambroise Paré - From Ambroise Paré To The Surgeon of The 21st Century, Or, From Surgical Ligation To RobotsDocument2 pagesAmbroise Paré - From Ambroise Paré To The Surgeon of The 21st Century, Or, From Surgical Ligation To Robotsorlandom61No ratings yet
- Ambroise Paré - DDocument1 pageAmbroise Paré - Dorlandom61No ratings yet
- Ambroise Paré (1510-1590) - On The Diagnosis and Treatment of Shoulder Dislocations - Markatos 2017Document4 pagesAmbroise Paré (1510-1590) - On The Diagnosis and Treatment of Shoulder Dislocations - Markatos 2017orlandom61No ratings yet
- Previously Malgaigne's Biography Surgical: Kenneth KeeleDocument1 pagePreviously Malgaigne's Biography Surgical: Kenneth Keeleorlandom61No ratings yet
- Ambroise Paré (1510-1590) and His Innovative Work On The Treatment of War Injuries - Markatos2017Document4 pagesAmbroise Paré (1510-1590) and His Innovative Work On The Treatment of War Injuries - Markatos2017orlandom61No ratings yet
- Ambroise Pare - Barber Vascular SurgeonDocument4 pagesAmbroise Pare - Barber Vascular Surgeonorlandom61No ratings yet
- Ambroise Paré, Surgery, and ObstetricsDocument2 pagesAmbroise Paré, Surgery, and Obstetricsorlandom61No ratings yet
- Re: "The River: A Journey To The Source of HIV and AIDS": American Journal of Epidemiology July 2002Document3 pagesRe: "The River: A Journey To The Source of HIV and AIDS": American Journal of Epidemiology July 2002VVVVNo ratings yet
- Ambroise Paré - Labor Improbus Omnia Vincit'' Ambroise Pare and Sixteenth Century Child CareDocument5 pagesAmbroise Paré - Labor Improbus Omnia Vincit'' Ambroise Pare and Sixteenth Century Child Careorlandom61No ratings yet
- A National Medical Response To Crisis - The Legacy of World War II - Nejm-Agosto - 2020Document3 pagesA National Medical Response To Crisis - The Legacy of World War II - Nejm-Agosto - 2020orlandom61No ratings yet
- Ambroise Paré (1510-1590) and His Innovative Work On The Treatment of War Injuries - Markatos2017Document4 pagesAmbroise Paré (1510-1590) and His Innovative Work On The Treatment of War Injuries - Markatos2017orlandom61No ratings yet
- Ambroise Paré and The Arrest of HaemorrhageDocument3 pagesAmbroise Paré and The Arrest of Haemorrhageorlandom61No ratings yet
- Ambroise Paré - 510 - 1590 - and - The - Evolution - of - The.94Document2 pagesAmbroise Paré - 510 - 1590 - and - The - Evolution - of - The.94orlandom61No ratings yet
- Ambroise Paré (1510-1590) - The Gentle SurgeonDocument2 pagesAmbroise Paré (1510-1590) - The Gentle Surgeonorlandom61No ratings yet
- Allen George Debus, 16 August 1926-6 March 2009Document5 pagesAllen George Debus, 16 August 1926-6 March 2009orlandom61No ratings yet
- Ambroise Pare (1510-1590) - Surgeon and Obstetrician of The RenaissanceDocument2 pagesAmbroise Pare (1510-1590) - Surgeon and Obstetrician of The Renaissanceorlandom61No ratings yet
- Ijarbs 7Document15 pagesIjarbs 7amanmalako50No ratings yet
- Referências MicrobiologiaDocument2 pagesReferências MicrobiologiaDanilo FerrazNo ratings yet
- DMJ 2020 0258Document17 pagesDMJ 2020 0258Elena Borş MorariNo ratings yet
- Total Knee Replacement: Done By: Sajeda Moha'dDocument27 pagesTotal Knee Replacement: Done By: Sajeda Moha'dlolo223No ratings yet
- Head To Toe AssessmentDocument2 pagesHead To Toe AssessmentAyucahyaNo ratings yet
- NP5Document12 pagesNP5rexinne noahNo ratings yet
- l8 Mock Test Pdce IIIDocument3 pagesl8 Mock Test Pdce IIINai IsaurraldeNo ratings yet
- National Health Policy and PlanDocument92 pagesNational Health Policy and PlanChetan BhattaNo ratings yet
- Ped A Internship 1517300917 PDF 2Document13 pagesPed A Internship 1517300917 PDF 2dariasuslowaNo ratings yet
- ANCA Associated Vasculitis After COVID 19Document7 pagesANCA Associated Vasculitis After COVID 19Arief KamilNo ratings yet
- Cardio Exercise Testing in Children and AdolecentsDocument288 pagesCardio Exercise Testing in Children and AdolecentsTeoNo ratings yet
- 2020 Pathology Compilation Questions by TopicsDocument7 pages2020 Pathology Compilation Questions by Topicsrupertgrint2000No ratings yet
- Microbiology Research ProposalDocument5 pagesMicrobiology Research Proposalapi-547677741No ratings yet
- Emphysema - Pleurisy - Pleural EffusionDocument17 pagesEmphysema - Pleurisy - Pleural EffusionGEN ERIGBUAGASNo ratings yet
- Nursing Diagnosis for TonsillitisDocument3 pagesNursing Diagnosis for TonsillitisVaneca Go67% (9)
- Rash Contacts Viral and Zoonotic VSN 2NHSCT171127Document69 pagesRash Contacts Viral and Zoonotic VSN 2NHSCT171127ana mariaNo ratings yet
- MCQ ParasitologyDocument7 pagesMCQ ParasitologyMontassar Dridi100% (1)
- Understanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionDocument27 pagesUnderstanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionNikhil BijuNo ratings yet
- Lecture Notes in ParasitologyDocument15 pagesLecture Notes in Parasitologyjiday23100% (5)
- Infectious Bronchitis in Broiler - Veterinaria DigitalDocument3 pagesInfectious Bronchitis in Broiler - Veterinaria DigitalMahesh KajagarNo ratings yet
- Noncardiogenic Pulmonary EdemaDocument12 pagesNoncardiogenic Pulmonary EdemaIrina DuceacNo ratings yet
- Rural MarketingDocument27 pagesRural MarketingnishantchopraNo ratings yet
- Health Shield 360 Enrollment FormDocument3 pagesHealth Shield 360 Enrollment FormgauravpassionNo ratings yet
- Beta Blocker TitrationDocument2 pagesBeta Blocker TitrationAwalunisa IstiqomahNo ratings yet
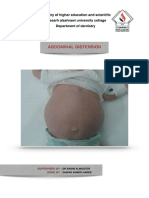
- Abdominal Distension: Supervised BY: Done BYDocument4 pagesAbdominal Distension: Supervised BY: Done BYŽäíñäb ÄljaÑabìNo ratings yet
- English-10 Q2-Module 5Document18 pagesEnglish-10 Q2-Module 5janeNo ratings yet
- Registering As A Dentist With The General Dental Council (Overseas Qualified)Document18 pagesRegistering As A Dentist With The General Dental Council (Overseas Qualified)zahra seraj0% (1)
- IMRAD pr1 Group 9Document7 pagesIMRAD pr1 Group 9ibanezlj30No ratings yet
- Microbiological Profile and Antimicrobial Susceptibility Pattern Ofaural Swab in Tertiary Care Hospital of Northern IndiaDocument7 pagesMicrobiological Profile and Antimicrobial Susceptibility Pattern Ofaural Swab in Tertiary Care Hospital of Northern IndiaIJAR JOURNALNo ratings yet
- Epstein-Barr Virus (Infectious Mononucleosis)Document3 pagesEpstein-Barr Virus (Infectious Mononucleosis)KanayaNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)