You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Medical Genetics at GlanceDocument116 pagesMedical Genetics at Glancedrabdulla977100% (4)
- Trial Different Mixing TimesDocument10 pagesTrial Different Mixing Timesrizky rajabillahNo ratings yet
- Trial Efficacy Dinotefuron To Control Whitefly in NurseryDocument4 pagesTrial Efficacy Dinotefuron To Control Whitefly in Nurseryrizky rajabillahNo ratings yet
- Disease Progression in Plantations of Acacia Mangium A Ffected by Red Root Rot (Ganoderma Philippii)Document14 pagesDisease Progression in Plantations of Acacia Mangium A Ffected by Red Root Rot (Ganoderma Philippii)rizky rajabillahNo ratings yet
- Chapter Three: TermitesDocument13 pagesChapter Three: Termitesrizky rajabillahNo ratings yet
- Morphological Character of Phlebiopsis sp.1 Isolates, A Potential Biological Control For Ganoderma PhilippiiDocument12 pagesMorphological Character of Phlebiopsis sp.1 Isolates, A Potential Biological Control For Ganoderma Philippiirizky rajabillahNo ratings yet
- Biodegradation Study of Pycnoporus Sanguineus and Its Effects On Structural and Chemical Features On Oil Palm Biomass ChipsDocument19 pagesBiodegradation Study of Pycnoporus Sanguineus and Its Effects On Structural and Chemical Features On Oil Palm Biomass Chipsrizky rajabillahNo ratings yet
- pr124 PDF 16794Document92 pagespr124 PDF 16794rizky rajabillahNo ratings yet
- Ketahanan Semai Akasia (Acacia Mangium) Pada Variasi Umur TERHADAP INFEKSI Ganoderma SPPDocument6 pagesKetahanan Semai Akasia (Acacia Mangium) Pada Variasi Umur TERHADAP INFEKSI Ganoderma SPPrizky rajabillahNo ratings yet
- Oxolinic Acid MSDS: Section 1: Chemical Product and Company IdentificationDocument5 pagesOxolinic Acid MSDS: Section 1: Chemical Product and Company Identificationrizky rajabillahNo ratings yet
- WRINKLED1 Specifies The Regulatory Action of LEAFY COTYLEDON2 Towards Fatty Acid Metabolism During Seed Maturation in ArabidopsisDocument14 pagesWRINKLED1 Specifies The Regulatory Action of LEAFY COTYLEDON2 Towards Fatty Acid Metabolism During Seed Maturation in Arabidopsisrizky rajabillahNo ratings yet
- Plant Unsaturated Fatty Acids: Biosynthesis and Regulation: Mei He, Chun-Xue Qin, Xu Wang and Nai-Zheng DingDocument13 pagesPlant Unsaturated Fatty Acids: Biosynthesis and Regulation: Mei He, Chun-Xue Qin, Xu Wang and Nai-Zheng Dingrizky rajabillahNo ratings yet
- 191-Article Text-655-9-10-20160202Document8 pages191-Article Text-655-9-10-20160202rizky rajabillahNo ratings yet
- 841-Article Text-16063-1-10-20201022Document7 pages841-Article Text-16063-1-10-20201022rizky rajabillahNo ratings yet
- The Effectiviness of Organic Method For Increasing The Productivity of Oil Palm That Is Infected by GanodermaDocument11 pagesThe Effectiviness of Organic Method For Increasing The Productivity of Oil Palm That Is Infected by Ganodermarizky rajabillahNo ratings yet
- Stem Cell Research ThesisDocument5 pagesStem Cell Research Thesisxdkankjbf100% (1)
- ANACARDIC CompressedDocument18 pagesANACARDIC CompressedElvaNo ratings yet
- Uploads571457146589cell Growth and Reproduction Key Study Guide PDFDocument12 pagesUploads571457146589cell Growth and Reproduction Key Study Guide PDFDiego GarciaNo ratings yet
- Adverse Event Following Immunization (AEFI) Manual of Procedures, 2014Document106 pagesAdverse Event Following Immunization (AEFI) Manual of Procedures, 2014Jezza Jonah AclanNo ratings yet
- 2.2. Digestion, Absorption, Secretion, and Utilization of Dietary LipidsDocument45 pages2.2. Digestion, Absorption, Secretion, and Utilization of Dietary LipidsAna AbuladzeNo ratings yet
- MCQ: Biophysics (Post Basic BSC Nursing)Document4 pagesMCQ: Biophysics (Post Basic BSC Nursing)varsha thakur100% (1)
- Exercises Genetics USTH2022Document15 pagesExercises Genetics USTH2022yungiang157No ratings yet
- First Detection of Bla TEM, SHV and CTX-M AmongDocument7 pagesFirst Detection of Bla TEM, SHV and CTX-M AmongHamidou OuedraogoNo ratings yet
- Hepatitis Virus: Pre Test and Post-TestDocument12 pagesHepatitis Virus: Pre Test and Post-TestDaffa IbnurasyNo ratings yet
- Compendium 2 ANSWERSDocument13 pagesCompendium 2 ANSWERSRajveen ChahalNo ratings yet
- PSGNDocument23 pagesPSGNalpha mae beltranNo ratings yet
- MCQ4FULLDocument139 pagesMCQ4FULLBruta Brutal100% (2)
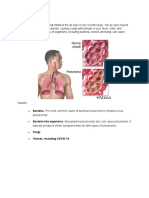
- Bacteria. The Most Common Cause of Bacterial Pneumonia Is StreptococcusDocument4 pagesBacteria. The Most Common Cause of Bacterial Pneumonia Is StreptococcusLouise OpinaNo ratings yet
- How To Improve Antifungal StewardshipDocument12 pagesHow To Improve Antifungal StewardshipciaranNo ratings yet
- Cell SignalingDocument53 pagesCell SignalingKaushiki KalravNo ratings yet
- Micropara 1Document8 pagesMicropara 1Jing DelfinNo ratings yet
- Advanced Drug Delivery Reviews: Erkki RuoslahtiDocument10 pagesAdvanced Drug Delivery Reviews: Erkki RuoslahtiacbgdvNo ratings yet
- STPM Trial 2009 Bio Q&A (Melaka)Document36 pagesSTPM Trial 2009 Bio Q&A (Melaka)SimPorNo ratings yet
- BACTE (Quizlet)Document4 pagesBACTE (Quizlet)Allyssa AniNo ratings yet
- HYPERSENTIVITYDocument4 pagesHYPERSENTIVITYVaishali PrasharNo ratings yet
- Xpert HIV-1 Qual: Faster Intervention For Better Patient CareDocument4 pagesXpert HIV-1 Qual: Faster Intervention For Better Patient CareKim LyNo ratings yet
- ImmunityDocument37 pagesImmunityRand Hussein100% (1)
- Introduction To MicrobiologyDocument5 pagesIntroduction To MicrobiologyChristina EscardaNo ratings yet
- Gizzard Proventriculus Health Marker Avinews Int Sept 22Document11 pagesGizzard Proventriculus Health Marker Avinews Int Sept 22mohamed helmyNo ratings yet
- Principles of Inheritance: Biohack NotesDocument9 pagesPrinciples of Inheritance: Biohack NotesRakshit ChaturvediNo ratings yet
- Sex LinkageDocument15 pagesSex LinkageWamia RahmanNo ratings yet
- Biology Chapter 15Document15 pagesBiology Chapter 15huifwiu jsbNo ratings yet
- Ii Pu Biology QPDocument6 pagesIi Pu Biology QPShambu BanavasiNo ratings yet
- Catalogue Kover SRL En.Document32 pagesCatalogue Kover SRL En.Ali Ben HusseinNo ratings yet