You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- JTlecture 01 PrintDocument17 pagesJTlecture 01 PrintJae-Soo ChangNo ratings yet
- 21 Golden Safety Rules. PortfolioDocument2 pages21 Golden Safety Rules. PortfolioPaul Bugeja100% (2)
- Hal Elrod & David Osborn & Honoree Corder - Miracle Morning Millionaires - What The Wealthy Do Before 8AM That Will Make You Rich (The Miracle Morning Book 11) (2018, Amazon Digital Services LLC) PDFDocument231 pagesHal Elrod & David Osborn & Honoree Corder - Miracle Morning Millionaires - What The Wealthy Do Before 8AM That Will Make You Rich (The Miracle Morning Book 11) (2018, Amazon Digital Services LLC) PDFtomycat100% (26)
- 3nursing Perspectives On The Impacts of COVID 19.2Document5 pages3nursing Perspectives On The Impacts of COVID 19.2SANo ratings yet
- Languages: Linguistic Diversity, Multilingualism, and Cognitive Skills: A Study of Disadvantaged Children in IndiaDocument22 pagesLanguages: Linguistic Diversity, Multilingualism, and Cognitive Skills: A Study of Disadvantaged Children in IndiaSANo ratings yet
- Beyond The Pandemic: COVID-19 Pandemic Changed The Face of LifeDocument20 pagesBeyond The Pandemic: COVID-19 Pandemic Changed The Face of LifeSANo ratings yet
- The Psychological and Social Impact of Covid-19: New Perspectives of Well-BeingDocument7 pagesThe Psychological and Social Impact of Covid-19: New Perspectives of Well-BeingSANo ratings yet
- A Research Agenda For The post-COVID-19 World: Theory and Research in Social PsychologyDocument8 pagesA Research Agenda For The post-COVID-19 World: Theory and Research in Social PsychologySANo ratings yet
- 1nihpp-2020 06 10 20127589Document10 pages1nihpp-2020 06 10 20127589SANo ratings yet
- Keraguard: Powerful Protection and Repair For HairDocument12 pagesKeraguard: Powerful Protection and Repair For HairSANo ratings yet
- Medicine: Social Distancing To Slow The US COVID-19 Epidemic: Longitudinal Pretest-Posttest Comparison Group StudyDocument12 pagesMedicine: Social Distancing To Slow The US COVID-19 Epidemic: Longitudinal Pretest-Posttest Comparison Group StudySANo ratings yet
- Social Isolation in Covid-19: The Impact of Loneliness: Debanjan Banerjee and Mayank RaiDocument3 pagesSocial Isolation in Covid-19: The Impact of Loneliness: Debanjan Banerjee and Mayank RaiSANo ratings yet
- Natural and Organic Cosmetics. Law Regulations and The Newest TrendsDocument7 pagesNatural and Organic Cosmetics. Law Regulations and The Newest TrendsSANo ratings yet
- Fig Latex Caprine Herpesvirus-1Document7 pagesFig Latex Caprine Herpesvirus-1SANo ratings yet
- Updated On: Descriptive SynopsisDocument3 pagesUpdated On: Descriptive SynopsisSANo ratings yet
- Lu 2020Document1 pageLu 2020SANo ratings yet
- Preparation of Herbello - An Herbal Antidandruff ShampooDocument9 pagesPreparation of Herbello - An Herbal Antidandruff ShampooSANo ratings yet
- Q&a Properties of The Hair and ScalpDocument5 pagesQ&a Properties of The Hair and ScalpSANo ratings yet
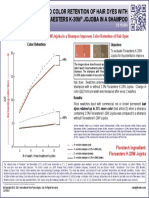
- Title Improved Color Retention of Hair Dyes With Floraesters K-20W Jojoba in A ShampooDocument1 pageTitle Improved Color Retention of Hair Dyes With Floraesters K-20W Jojoba in A ShampooSANo ratings yet
- Inhydrate: Moisturises From The Inside OutDocument1 pageInhydrate: Moisturises From The Inside OutSANo ratings yet
- Hydrate: Moisturises The Skin From The Inside OutDocument2 pagesHydrate: Moisturises The Skin From The Inside OutSANo ratings yet
- Vercare: Anionic Polyester-Polyurethane DispersionDocument4 pagesVercare: Anionic Polyester-Polyurethane DispersionSANo ratings yet
- En 1clr-7022 Cutipure CLRDocument7 pagesEn 1clr-7022 Cutipure CLRSANo ratings yet
- Invert: Fights Aging From The Inside OutDocument2 pagesInvert: Fights Aging From The Inside OutSANo ratings yet
- Safety Data Sheet: According To Regulation (EC) No. 1907/2006 (REACH)Document6 pagesSafety Data Sheet: According To Regulation (EC) No. 1907/2006 (REACH)SANo ratings yet
- A Natural Oil To Cure and To ProtectDocument2 pagesA Natural Oil To Cure and To ProtectSANo ratings yet
- Construction Management: Divya Negi Faculty, B.Voc., ABITDocument11 pagesConstruction Management: Divya Negi Faculty, B.Voc., ABITDIVYA NEGINo ratings yet
- Challenges and Solutions For Tunnel ConsDocument29 pagesChallenges and Solutions For Tunnel ConsAfomiya ZelalemNo ratings yet
- Adobe Illustrator CS5 Keyboard ShortcutsDocument9 pagesAdobe Illustrator CS5 Keyboard ShortcutsDara Maria0% (1)
- ESCAFEDocument10 pagesESCAFEBenz Joven BarbonNo ratings yet
- Medifa 8000 Service ManuelDocument44 pagesMedifa 8000 Service ManuelhoudaNo ratings yet
- Electric SignalsDocument29 pagesElectric Signalsyttan1116No ratings yet
- "Lemko Experiences As Recalled by Teodor Doklia" (Yasiunka and Other Villages)Document31 pages"Lemko Experiences As Recalled by Teodor Doklia" (Yasiunka and Other Villages)TheLemkoProjectNo ratings yet
- REEandRME0918se jg18Document116 pagesREEandRME0918se jg18TOPNOTCHER Philippines0% (1)
- N45UH Grade Neodymium Magnets DataDocument1 pageN45UH Grade Neodymium Magnets DataSteve HsuNo ratings yet
- Drill Stem TestDocument3 pagesDrill Stem Testpoliskarma0% (1)
- DT4 Student's Book - U3Document16 pagesDT4 Student's Book - U3Tuyên LươngNo ratings yet
- Design of EW CircuitDocument8 pagesDesign of EW Circuitpatrick.harris881No ratings yet
- Fuse Link SizeDocument14 pagesFuse Link Sizebilly12379224No ratings yet
- METU ENVE202 Laboratory ManualDocument47 pagesMETU ENVE202 Laboratory ManualSarp ÇelebiNo ratings yet
- Equipment List For 1 X 750 MW Gas Based Combined Cycle Power PlantDocument3 pagesEquipment List For 1 X 750 MW Gas Based Combined Cycle Power PlantRazi Khan100% (1)
- Inmetro 001Document15 pagesInmetro 001Anderson JuniorNo ratings yet
- NumeracyDocument12 pagesNumeracySerah BijuNo ratings yet
- Neets Project V GuardDocument81 pagesNeets Project V GuardNeetha Kanjakulath K67% (6)
- Comparing Successful and Unsuccessful Brand Positioning Strategies in Automobile SectorDocument3 pagesComparing Successful and Unsuccessful Brand Positioning Strategies in Automobile SectorRishabh ChoudharyNo ratings yet
- Aerscreen UserguideDocument104 pagesAerscreen UserguideAndres Muro ManriqueNo ratings yet
- CSD Okara (2) OriDocument1 pageCSD Okara (2) OriZeeshan ch 'Hadi'No ratings yet
- Seminar03 MPE TrussDocument6 pagesSeminar03 MPE TrussAndrei CioroianuNo ratings yet
- Canon Eos 300d Service ManualDocument109 pagesCanon Eos 300d Service ManualEnrique RichardNo ratings yet
- DNA and Proteins Doodle Notes 2Document10 pagesDNA and Proteins Doodle Notes 2Leila BeyyoudhNo ratings yet
- Cálculo Fecha Redención 5778 (2018) - Rav AshlagDocument176 pagesCálculo Fecha Redención 5778 (2018) - Rav AshlagDavid Saportas LièvanoNo ratings yet
- Mto Shear ScriptDocument7 pagesMto Shear ScriptAustria, Gerwin Iver LuisNo ratings yet