You might also like
- Khmer Friendship HospitalDocument3 pagesKhmer Friendship Hospitalខៃសិញ- KhaisinhNo ratings yet
- Listening A06Document5 pagesListening A06qartzz100% (1)
- TRADUCCION (1) 1 - The COVID-19 Epidemic PDFDocument3 pagesTRADUCCION (1) 1 - The COVID-19 Epidemic PDFSikamelNo ratings yet
- Mixed-Species Malaria Infections in HumansDocument8 pagesMixed-Species Malaria Infections in Humansdella rafika sariNo ratings yet
- Mixed Malaria Infection: About An Imported Case and Review of The LiteratureDocument5 pagesMixed Malaria Infection: About An Imported Case and Review of The LiteratureIJAR JOURNALNo ratings yet
- Clinical Profile of Plasmodium Falciparum andDocument11 pagesClinical Profile of Plasmodium Falciparum andCarlos Javier Castro CavadiaNo ratings yet
- University of Santo Tomas: UST-SHS Practical Research 2Document3 pagesUniversity of Santo Tomas: UST-SHS Practical Research 2MayNo ratings yet
- How Not To Miss A Case of Malaria in Emergency DepDocument17 pagesHow Not To Miss A Case of Malaria in Emergency Depnadila raudhaniNo ratings yet
- Severe Morbidity and Mortality Risk From Malaria in The United States, 1985 - 2011Document8 pagesSevere Morbidity and Mortality Risk From Malaria in The United States, 1985 - 2011Hermansyah ChiuNo ratings yet
- Malaria Case SeriesDocument3 pagesMalaria Case Seriesnancy voraNo ratings yet
- Malaria in IcuDocument13 pagesMalaria in Iculakshminivas PingaliNo ratings yet
- 2021 10 27 21265557 FullDocument22 pages2021 10 27 21265557 FullZasaNo ratings yet
- Tropicalmed 07 00260Document21 pagesTropicalmed 07 00260فرجني موغNo ratings yet
- (2014) Parasitic Pneumonia and Lung InvolvementDocument18 pages(2014) Parasitic Pneumonia and Lung InvolvementAlan DiazNo ratings yet
- JCDR MalariaDocument4 pagesJCDR Malariashruti kumarNo ratings yet
- RetrovirusesDocument7 pagesRetroviruseseluardpaul37No ratings yet
- Study of Association of Thrombocytopenia With Plasmodium Vivax Infection Other Infections Original ArticleDocument3 pagesStudy of Association of Thrombocytopenia With Plasmodium Vivax Infection Other Infections Original ArticleEnvhy AmaliaNo ratings yet
- Case Report Plasmodium Vivax Infected PatientDocument8 pagesCase Report Plasmodium Vivax Infected Patientjohanna monsalveNo ratings yet
- MPV 12Document6 pagesMPV 12Tetteh JudeNo ratings yet
- Antiviral Nucleosides-Flu Viruses-Quinolines-COVID-19: Corresponding AuthorDocument12 pagesAntiviral Nucleosides-Flu Viruses-Quinolines-COVID-19: Corresponding AuthorDrkrishnasarma pathyNo ratings yet
- Haq2021 Article SARS CoV 2BigSeroprevalenceDatDocument6 pagesHaq2021 Article SARS CoV 2BigSeroprevalenceDatMuhammad Asif NaveedNo ratings yet
- 05 - Approfondimenti MalariaDocument7 pages05 - Approfondimenti Malariamy accountNo ratings yet
- Prevalence and Intensity of Loa Loa Infection Over Twenty-Three Years in Three Communities of The Mbalmayo Health District (Central Cameroon)Document7 pagesPrevalence and Intensity of Loa Loa Infection Over Twenty-Three Years in Three Communities of The Mbalmayo Health District (Central Cameroon)Alyaa IrawanNo ratings yet
- Kumpulan Jurnal DengueDocument122 pagesKumpulan Jurnal DengueMas MantriNo ratings yet
- Repassmalaria 2Document6 pagesRepassmalaria 2api-329029118No ratings yet
- Summarization of The ArticleDocument2 pagesSummarization of The ArticleHershei Vonne BaccayNo ratings yet
- Parasit Pipi PDFDocument15 pagesParasit Pipi PDFRNo ratings yet
- Influenzaandviral Pneumonia: Rodrigo Cavallazzi,, Julio A. RamirezDocument19 pagesInfluenzaandviral Pneumonia: Rodrigo Cavallazzi,, Julio A. Ramirezlevani memarneNo ratings yet
- INTRODUCTIONDocument2 pagesINTRODUCTIONhumamhamodyNo ratings yet
- Malaria: What's New in The Management of Malaria?Document22 pagesMalaria: What's New in The Management of Malaria?wiwiNo ratings yet
- New MJAFIDocument5 pagesNew MJAFICI BorderprotectionNo ratings yet
- Common Invasive Fungal Diseases An Overview of Invasive Candidiasis, Aspergillosis, Cryptococcosis, and Pneumocystis PneumoniaDocument12 pagesCommon Invasive Fungal Diseases An Overview of Invasive Candidiasis, Aspergillosis, Cryptococcosis, and Pneumocystis PneumoniaFebri Dwi HaryonoNo ratings yet
- Concurrent Malaria and Typhoid Fever in The Tropics: The Diagnostic Challenges and Public Health ImplicationsDocument10 pagesConcurrent Malaria and Typhoid Fever in The Tropics: The Diagnostic Challenges and Public Health Implicationselsa_imamNo ratings yet
- 2022 Article 775Document17 pages2022 Article 775Felipe CortesNo ratings yet
- Spatial Distribution of G6PD Deficiency Variants Across Malaria-Endemic RegionsDocument15 pagesSpatial Distribution of G6PD Deficiency Variants Across Malaria-Endemic RegionsdesyNo ratings yet
- A Study of Thrombocytopenia in Malaria and Its Prognostic SignificanceDocument6 pagesA Study of Thrombocytopenia in Malaria and Its Prognostic SignificanceBobby Faisyal RakhmanNo ratings yet
- Velavan Et Al-2020-Tropical Medicine International HealthDocument4 pagesVelavan Et Al-2020-Tropical Medicine International Healthjunior rodriguesNo ratings yet
- Serious Fungal Infections in PakistanDocument8 pagesSerious Fungal Infections in PakistanMuhammad Vaqar HussainNo ratings yet
- Viruses 09 00011Document17 pagesViruses 09 00011Hany ZutanNo ratings yet
- Antimalarial Drugs: Modes of Action and Mechanisms of Parasite ResistanceDocument18 pagesAntimalarial Drugs: Modes of Action and Mechanisms of Parasite ResistancePuspa Lestari RyathNo ratings yet
- Clinical Immunology: Susanna Felsenstein, Jenny A. Herbert, Paul S. Mcnamara, Christian M. Hedrich TDocument13 pagesClinical Immunology: Susanna Felsenstein, Jenny A. Herbert, Paul S. Mcnamara, Christian M. Hedrich TIrma NeyraNo ratings yet
- DualtherapeuticapproachforCOVID 19 Draft Noa 1Document10 pagesDualtherapeuticapproachforCOVID 19 Draft Noa 1ohlakhNo ratings yet
- CD14CD16 PDFDocument16 pagesCD14CD16 PDFAde OktiviyariNo ratings yet
- UntitledDocument82 pagesUntitled乙骨憂太No ratings yet
- Covid EditorialDocument10 pagesCovid EditorialtersiaNo ratings yet
- Morphological Features and Differential Counts of Plasmodium Knowlesi Parasites in Naturally Acquired Human InfectionsDocument10 pagesMorphological Features and Differential Counts of Plasmodium Knowlesi Parasites in Naturally Acquired Human InfectionsAnantaBenvenutoNo ratings yet
- The Prevention and Treatment of Plasmodium: Vivax MalariaDocument21 pagesThe Prevention and Treatment of Plasmodium: Vivax MalariaDoaa SaifNo ratings yet
- Im Wong 2011Document12 pagesIm Wong 2011ibanggNo ratings yet
- Articulo Revision Vih 04.10Document10 pagesArticulo Revision Vih 04.10Enrique MeccaNo ratings yet
- The Prevalence of Parasitic Protozoan Diseases in Iraq, 2016Document5 pagesThe Prevalence of Parasitic Protozoan Diseases in Iraq, 2016Jeremia AnkesaNo ratings yet
- Kaposi SarcomaDocument11 pagesKaposi Sarcomaedgar mandengNo ratings yet
- Nutrients: The Antiviral, Anti-Inflammatory E Medicinal Herbs and Mushrooms and Sars-Cov-2 InfectionDocument13 pagesNutrients: The Antiviral, Anti-Inflammatory E Medicinal Herbs and Mushrooms and Sars-Cov-2 Infectionmmbire@gmail.comNo ratings yet
- WholeDocument3 pagesWholePQ TODANo ratings yet
- 8 Malaria Is An Uncommon Cause of Adult Sepsis in SouthDocument9 pages8 Malaria Is An Uncommon Cause of Adult Sepsis in SouthAhmad Zaki YamaniNo ratings yet
- MalariaDocument19 pagesMalariamohanNo ratings yet
- A Comprehensive Summary of The Knowledge On COVID-19 TreatmentDocument37 pagesA Comprehensive Summary of The Knowledge On COVID-19 TreatmentChawki MokademNo ratings yet
- Infections Diseases MRCP Crib Sheet by Law and MedicineDocument33 pagesInfections Diseases MRCP Crib Sheet by Law and MedicineMatin Ahmad KhanNo ratings yet
- RSVDocument8 pagesRSVilyas9558No ratings yet
- Diagnosis of Leishmaniasis: Review ArticleDocument12 pagesDiagnosis of Leishmaniasis: Review ArticleTUfoodbiotech groupNo ratings yet
- Leptospirosis: Tropical To Subtropical India: EditorialDocument2 pagesLeptospirosis: Tropical To Subtropical India: EditorialTiara AnggrainiNo ratings yet
- UntitledDocument26 pagesUntitledLaura SanchezNo ratings yet
- What Is Reye's SyndromeDocument3 pagesWhat Is Reye's SyndromeRanvabesa Cakra Laksa CitaNo ratings yet
- Nursing Process Focus: Patients Receiving Fluticasone (Flonase) Assessment Potential Nursing DiagnosesDocument2 pagesNursing Process Focus: Patients Receiving Fluticasone (Flonase) Assessment Potential Nursing DiagnosesHannah Lorraine GamayonNo ratings yet
- CellulitisDocument5 pagesCellulitisKrishna PendemNo ratings yet
- Sta. Ana Hospital: Covid-19 Testing LaboratoryDocument1 pageSta. Ana Hospital: Covid-19 Testing LaboratoryRamel Yen CerantesNo ratings yet
- Brain Abscess: Mohd Roslee Bin Abd GhaniDocument36 pagesBrain Abscess: Mohd Roslee Bin Abd GhaniSaha DirllahNo ratings yet
- Influenza Nausea and Vomiting FeverDocument5 pagesInfluenza Nausea and Vomiting Feverjayne_saulogNo ratings yet
- Ministry of HealthDocument7 pagesMinistry of Healthks jNo ratings yet
- The Major Health Implicationsof Social ConnectionDocument9 pagesThe Major Health Implicationsof Social ConnectionMaggay LarsNo ratings yet
- Ulcerativecolitis 170323180448 PDFDocument88 pagesUlcerativecolitis 170323180448 PDFBasudewo Agung100% (1)
- Reading Sub-Test: Answer Key - Part ADocument24 pagesReading Sub-Test: Answer Key - Part AAlwin BrightNo ratings yet
- Sankalak - 2023 Estimates - NACODocument372 pagesSankalak - 2023 Estimates - NACOjeancsixNo ratings yet
- ZMK SchizophreniaDocument73 pagesZMK SchizophreniaAhlam Azam, MohamedNo ratings yet
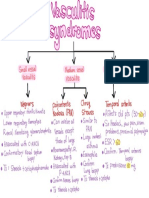
- Vasculitis SyndromesDocument1 pageVasculitis SyndromesamanNo ratings yet
- Altered Body TemperatureDocument27 pagesAltered Body TemperatureAnusikta PandaNo ratings yet
- 23 08 21Document3 pages23 08 21JENNIFER JOHN MBBS2020No ratings yet
- Brochure - PregnancyDocument3 pagesBrochure - PregnancyhiyceNo ratings yet
- Virology MCQsDocument7 pagesVirology MCQsHabib Ullah100% (1)
- Armodafinil and ModafinilDocument1 pageArmodafinil and ModafinilJayson TribacoNo ratings yet
- Content CDC and MDDocument7 pagesContent CDC and MDAmir Ahmed GezaNo ratings yet
- From Rare Diseases, Genetic Disorders To Orphan DrugsDocument192 pagesFrom Rare Diseases, Genetic Disorders To Orphan DrugsShing Ming TangNo ratings yet
- English Hand BookDocument55 pagesEnglish Hand BookVeena Jayaram Rao RajgopalNo ratings yet
- Lesson PlanDocument8 pagesLesson PlanUpendra Nayka50% (2)
- Obana ProtocolDocument4 pagesObana Protocolpaiskopaxiew888No ratings yet
- Inpatient Asthma: Clinical PathwayDocument9 pagesInpatient Asthma: Clinical PathwaydedeNo ratings yet
- Exposé VIHDocument6 pagesExposé VIHWilfried TougmaNo ratings yet
- Rifemedic 7 Program List 2020Document44 pagesRifemedic 7 Program List 2020pierre van wykNo ratings yet
- Case PresentationDocument13 pagesCase PresentationAnto BijuNo ratings yet
- PTS Ganjil Kelas 10 LMDocument7 pagesPTS Ganjil Kelas 10 LMAulia FatihaNo ratings yet