You might also like
- Isolation and Characterization of Endophytic Actinomycetes from Mangrove PlantsDocument12 pagesIsolation and Characterization of Endophytic Actinomycetes from Mangrove PlantsFebry OploverzNo ratings yet
- Zenniel Et. Al., 2002Document13 pagesZenniel Et. Al., 2002Aswawi AwiNo ratings yet
- 2012 Chandra JPPRDocument6 pages2012 Chandra JPPRknchandu1No ratings yet
- Transfer of Salmonella Enterica Serovar Typhimurium From Contaminated Irrigation Water To Parsley Is Dependent On Curli and Cellulose, The Biofilm Matrix ComponentsDocument6 pagesTransfer of Salmonella Enterica Serovar Typhimurium From Contaminated Irrigation Water To Parsley Is Dependent On Curli and Cellulose, The Biofilm Matrix ComponentshugoNo ratings yet
- 2198.full 2Document11 pages2198.full 2Maya SariNo ratings yet
- Pythium Species Associated With Rice Stand Establishment Problems in ArkansasDocument7 pagesPythium Species Associated With Rice Stand Establishment Problems in ArkansasSadao MatsumotoNo ratings yet
- Isolation and Identification of Antagonistic Bacterium Against Pathogens of Bacterial Tuber Rot of Amorphophallus MuelleriDocument8 pagesIsolation and Identification of Antagonistic Bacterium Against Pathogens of Bacterial Tuber Rot of Amorphophallus MuelleriRuby SakaNo ratings yet
- Evaluation of Phosphate Solubilizing Bacteria Isolated From Rhizospheric Soil of Parthenium PlantDocument7 pagesEvaluation of Phosphate Solubilizing Bacteria Isolated From Rhizospheric Soil of Parthenium PlantJournal of Biotechnology and Crop SciencesNo ratings yet
- In Vitro Efficacy of Entomopathogenic Nematodes (EPNS) Against Economically Important Insect-Pests of CauliflowerDocument9 pagesIn Vitro Efficacy of Entomopathogenic Nematodes (EPNS) Against Economically Important Insect-Pests of CauliflowerMamta AgarwalNo ratings yet
- Ajabssp 2009 146 151Document6 pagesAjabssp 2009 146 151Hilma Dianti MarhamNo ratings yet
- 10.1515 - ZNC 2009 5 618Document6 pages10.1515 - ZNC 2009 5 618Sona HaciliNo ratings yet
- 30 36 PDFDocument7 pages30 36 PDFray m deraniaNo ratings yet
- International Journal of PharmacyDocument5 pagesInternational Journal of PharmacyVardhana JanakiramanNo ratings yet
- Appl. Environ. Microbiol.-2005-Compant-1685-93Document9 pagesAppl. Environ. Microbiol.-2005-Compant-1685-93Lucas Antonio GallaratoNo ratings yet
- Báo-Invitro Inhibition of Growth of Some Seedling Blight Inducing Pathogens by Compost-Inhabiting MicrobesDocument4 pagesBáo-Invitro Inhibition of Growth of Some Seedling Blight Inducing Pathogens by Compost-Inhabiting MicrobesYến LinhNo ratings yet
- Identification of Leaf Spot Causing Pathogen Through ITS Sequencing in Clove and It's Management Using Bioagents, Botanicals and ChemicalsDocument7 pagesIdentification of Leaf Spot Causing Pathogen Through ITS Sequencing in Clove and It's Management Using Bioagents, Botanicals and ChemicalsEditor IJTSRDNo ratings yet
- Isolation of RhizosphereDocument7 pagesIsolation of RhizosphereAnisam AbhiNo ratings yet
- Isolation and Identification of Streptomyces From Different Sample of SoilsDocument6 pagesIsolation and Identification of Streptomyces From Different Sample of SoilsMandy FisherNo ratings yet
- AJOL Journal File 1Document10 pagesAJOL Journal File 1GeeVee23No ratings yet
- Corms as a source of explants for the successful clonal propagation of Crocus cancellatusDocument5 pagesCorms as a source of explants for the successful clonal propagation of Crocus cancellatusvivekabarathNo ratings yet
- 22 PDFDocument6 pages22 PDFParishay BatoolNo ratings yet
- A Systematic Review On Isolation, Identification, and Characterization of Agrobacterium Tumefaciens From Leguminous PlantsDocument5 pagesA Systematic Review On Isolation, Identification, and Characterization of Agrobacterium Tumefaciens From Leguminous PlantsMiddle East Journal of Applied Science & TechnologyNo ratings yet
- Pdis 2002 86 2 127Document4 pagesPdis 2002 86 2 127Aspin PianNo ratings yet
- Isolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological FeaturesDocument7 pagesIsolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological Featurescrashoverride29187No ratings yet
- Chen1997 Article SuccessfulInterspecificHybridiDocument7 pagesChen1997 Article SuccessfulInterspecificHybridiRifqi ZauhairNo ratings yet
- Study o F Callus Tissues From Different Parts of S. S. C. Roy 1Document2 pagesStudy o F Callus Tissues From Different Parts of S. S. C. Roy 1Alex CriolloNo ratings yet
- Theobroma Cacao PhytophthoraDocument10 pagesTheobroma Cacao PhytophthoraOpenaccess Research paperNo ratings yet
- Isolation and Characterization of Bioactive Actinomycetes From Soil in and Around NagpurDocument6 pagesIsolation and Characterization of Bioactive Actinomycetes From Soil in and Around NagpurAjzm CompanyNo ratings yet
- 24 Vol. 4 Issue 4 IJPSR April 2013 RA 2207 Paper 24Document6 pages24 Vol. 4 Issue 4 IJPSR April 2013 RA 2207 Paper 24Ajzm CompanyNo ratings yet
- M.P.prasad and Sunayana DagarDocument11 pagesM.P.prasad and Sunayana DagarAbhishek SinghNo ratings yet
- Rhizobium Meliloti and Their Biocontrol PotentialDocument5 pagesRhizobium Meliloti and Their Biocontrol PotentialYogesh VishwakarmaNo ratings yet
- Jurnal Mycorrhiza-05-017Document6 pagesJurnal Mycorrhiza-05-017Azhari RizalNo ratings yet
- Antifungal Potential of Extracellular Metabolites Produced by StreptomyDocument8 pagesAntifungal Potential of Extracellular Metabolites Produced by Streptomysiddhipurohit134586No ratings yet
- Clitoria Ternatea in Thailand: Isolation and Characterization of Rhizobia From Nodules ofDocument7 pagesClitoria Ternatea in Thailand: Isolation and Characterization of Rhizobia From Nodules ofVeimer ChanNo ratings yet
- 38 Ijasrapr201738Document6 pages38 Ijasrapr201738TJPRC PublicationsNo ratings yet
- Merging Biotechnology With Biological Control: Banana Musa Tissue Culture Plants Enhanced by Endophytic FungiDocument7 pagesMerging Biotechnology With Biological Control: Banana Musa Tissue Culture Plants Enhanced by Endophytic FungiAkash DoiphodeNo ratings yet
- J Mimet 2005 04 031Document7 pagesJ Mimet 2005 04 031indra maulanaNo ratings yet
- 26-04-2021-1619434181-8-Ijans-3. Ijans - Stored Pulses-Isolation of Fungi in Special Reference With Green, Red, Black Grams and MassorDocument8 pages26-04-2021-1619434181-8-Ijans-3. Ijans - Stored Pulses-Isolation of Fungi in Special Reference With Green, Red, Black Grams and Massoriaset123No ratings yet
- Detection of Alternaria radicina and A. dauci from Imported Carrot SeedDocument7 pagesDetection of Alternaria radicina and A. dauci from Imported Carrot SeedPaula Ordenes DomínguezNo ratings yet
- Phenotypic Molecular and Biochemical Evaluation ofDocument11 pagesPhenotypic Molecular and Biochemical Evaluation ofAmina BelguendouzNo ratings yet
- Black spot disease resistance in carrotsDocument2 pagesBlack spot disease resistance in carrotsBongile NcantswaNo ratings yet
- Anthracnose Pathogen of The Succulent Plant Echeveria Perle Von NürnbergDocument4 pagesAnthracnose Pathogen of The Succulent Plant Echeveria Perle Von NürnbergJosue RomanNo ratings yet
- Phaseolus Vulgarisseed-Borne Endophytic Community With Novel Bacterial Species Such As Rhizobium Endophyticum Sp. Nov.Document6 pagesPhaseolus Vulgarisseed-Borne Endophytic Community With Novel Bacterial Species Such As Rhizobium Endophyticum Sp. Nov.lesousa454No ratings yet
- 2011 Article 205Document8 pages2011 Article 205Hanylu AvilaNo ratings yet
- Brassica Oleracea Capitata L. Pratylenchus PenetransDocument7 pagesBrassica Oleracea Capitata L. Pratylenchus PenetransHalimah SiregarNo ratings yet
- Evaluation of Bacterial Antagonists of Ralstonia Solanacearum, CausalDocument6 pagesEvaluation of Bacterial Antagonists of Ralstonia Solanacearum, CausalHvzem MohammedNo ratings yet
- Potential Impact of Alternaria Macrospora On Cotton Production in West TexasDocument14 pagesPotential Impact of Alternaria Macrospora On Cotton Production in West TexasStephen MooreNo ratings yet
- Isolation and Identification of Antibiotic-Producing Actinom YcetesDocument8 pagesIsolation and Identification of Antibiotic-Producing Actinom YcetesrababNo ratings yet
- Bacterial and Viral Pathogens AssociatedDocument10 pagesBacterial and Viral Pathogens Associatednanochabow537No ratings yet
- Antimicrobial Properties of Vermicompost Associated MycofloraDocument4 pagesAntimicrobial Properties of Vermicompost Associated Mycofloraseematj2617No ratings yet
- Published Paper 11-7 - 6025 - PDFDocument11 pagesPublished Paper 11-7 - 6025 - PDFAhmed IsmailNo ratings yet
- 2013.18.endophytic Mycobiota Characterization of The AmazonianDocument5 pages2013.18.endophytic Mycobiota Characterization of The AmazonianfeNo ratings yet
- Biological Management of Groundnut Stemrot Pathogen: Sclerotium Rolfsii (Sacc.) by TrichodermavirideDocument8 pagesBiological Management of Groundnut Stemrot Pathogen: Sclerotium Rolfsii (Sacc.) by TrichodermavirideTJPRC PublicationsNo ratings yet
- Sequence Related Amplified Polymorphism (SRAP) Marker, Potato, Bacterial Wilt.Document9 pagesSequence Related Amplified Polymorphism (SRAP) Marker, Potato, Bacterial Wilt.Linh NguyenNo ratings yet
- Mycorrhiza 05 053Document9 pagesMycorrhiza 05 053Azhari RizalNo ratings yet
- Technique For Screening of Apple and Pear Germplasm Against White Root Rot (Dematophora Necatrix)Document3 pagesTechnique For Screening of Apple and Pear Germplasm Against White Root Rot (Dematophora Necatrix)Shailendra RajanNo ratings yet
- Cajanol DetectionDocument8 pagesCajanol DetectionDrJohn Milton LimaNo ratings yet
- Isolation and Screening of Actinomycetes From Sundarbans Soil For Antibacterial ActivityDocument5 pagesIsolation and Screening of Actinomycetes From Sundarbans Soil For Antibacterial Activityanggi marlianaNo ratings yet
- Soil Fungi and Soil Fertility: An Introduction to Soil MycologyFrom EverandSoil Fungi and Soil Fertility: An Introduction to Soil MycologyRating: 3.5 out of 5 stars3.5/5 (2)
- Caelus TT - Caelus Product TrainingDocument93 pagesCaelus TT - Caelus Product TrainingYader Daniel Ortiz CerdaNo ratings yet
- Capacitores Electroliticos SMDDocument8 pagesCapacitores Electroliticos SMDalvaro marrugoNo ratings yet
- AstigmatismDocument1 pageAstigmatismAmmellya PutriNo ratings yet
- Common Carrier RulingDocument4 pagesCommon Carrier RulingChic PabalanNo ratings yet
- Assignment 12Document7 pagesAssignment 12Anonymous mqIqN5zNo ratings yet
- Ahmed Ali Alshehri Mobile 00966-508-217284 Address: P.O.Box 70132 Zipcode:31952 Riyadh Street, Al-Khobar, Saudi ArabiaDocument9 pagesAhmed Ali Alshehri Mobile 00966-508-217284 Address: P.O.Box 70132 Zipcode:31952 Riyadh Street, Al-Khobar, Saudi ArabiaShah KhalidNo ratings yet
- 22 Imobilisasi Pada Usia LanjutDocument34 pages22 Imobilisasi Pada Usia LanjutGian KalalembangNo ratings yet
- TheologicalDiscipleship English 2011-1Document144 pagesTheologicalDiscipleship English 2011-1bilnyangnatNo ratings yet
- MSDS 442 BulkDocument2 pagesMSDS 442 BulkEliasNo ratings yet
- 16 Balance TestDocument15 pages16 Balance Testelga saniNo ratings yet
- Siemens 810dDocument13 pagesSiemens 810dAdo ŠehićNo ratings yet
- Phychem Chapter 3 Part 1Document14 pagesPhychem Chapter 3 Part 1Skye DiazNo ratings yet
- Material Safety Data Sheet: Chemtherm 550Document6 pagesMaterial Safety Data Sheet: Chemtherm 550Javier vaca villalbaNo ratings yet
- El Anatsui - TransformationsDocument15 pagesEl Anatsui - TransformationsReece BriceNo ratings yet
- ZTE UMTS KPI Optimization Analysis Guide V1 1 1Document62 pagesZTE UMTS KPI Optimization Analysis Guide V1 1 1GetitoutLetitgo100% (1)
- Singaporean Notices To Mariners: Section ContentDocument35 pagesSingaporean Notices To Mariners: Section ContentGaurav SoodNo ratings yet
- Revision 2 Pages Grammar Drills 106078Document3 pagesRevision 2 Pages Grammar Drills 106078Ligia Dragomir100% (1)
- ISO 14000 - WikipediaDocument5 pagesISO 14000 - WikipediaalexokorieNo ratings yet
- Worthy - Elevation WorshipDocument1 pageWorthy - Elevation WorshipSabine J.100% (1)
- Matrox DSX Le4Document76 pagesMatrox DSX Le4doc docNo ratings yet
- Shadows On The Moon by Zoe Marriott ExtractDocument20 pagesShadows On The Moon by Zoe Marriott ExtractWalker Books100% (1)
- Israel Is The MessIahDocument288 pagesIsrael Is The MessIahTeerayoot NoidaNo ratings yet
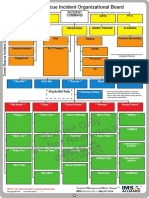
- Sample Only Do Not Reproduce: Trench Rescue Incident Organizational BoardDocument1 pageSample Only Do Not Reproduce: Trench Rescue Incident Organizational BoardLuis Diaz CerdanNo ratings yet
- MayankDocument38 pagesMayankmayank13430No ratings yet
- Water Spray Nozzle: Fire Fighting Equipment Data SheetDocument7 pagesWater Spray Nozzle: Fire Fighting Equipment Data SheetJosef MadronaNo ratings yet
- Bronchiolitis A Practical Approach For The General RadiologistDocument42 pagesBronchiolitis A Practical Approach For The General RadiologistTara NareswariNo ratings yet
- Impex Kitchen Appliances PDFDocument4 pagesImpex Kitchen Appliances PDFqwqw11No ratings yet
- Porsche Boxster Versus Factory Five 818Document8 pagesPorsche Boxster Versus Factory Five 818SteveNo ratings yet
- Philips HD5 enDocument5 pagesPhilips HD5 enmohamed boufasNo ratings yet
- Rental Range Powered by YANMAR Diesel GeneratorDocument6 pagesRental Range Powered by YANMAR Diesel GeneratorJonathan CastilloNo ratings yet