You might also like
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5813)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Awesome Review 2018 Internal Medicine ABIM Board Review (PDFDrive)Document592 pagesAwesome Review 2018 Internal Medicine ABIM Board Review (PDFDrive)saideh naghibiNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Uworld Peds MicroDocument5 pagesUworld Peds MicroJoan ChoiNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Typhoid FeverDocument3 pagesTyphoid Feverdejay100No ratings yet
- Kaposi SarcomaDocument41 pagesKaposi SarcomaMihaela Pal0% (1)
- Virology Lab Who GuidelinesDocument58 pagesVirology Lab Who GuidelinesIkhlasNo ratings yet
- Dental Considerations For Patients With Lupus ErythematosusDocument55 pagesDental Considerations For Patients With Lupus ErythematosusPhyo Ko KoNo ratings yet
- CestodaDocument57 pagesCestodaJingga TuasamuNo ratings yet
- RotavirusDocument6 pagesRotavirusRevant AgarwalNo ratings yet
- Viral Hepatitis OverviewDocument22 pagesViral Hepatitis OverviewNifemi OnatoyeNo ratings yet
- Acute Lymphoblastic Leukemia: Differential DiagnosisDocument6 pagesAcute Lymphoblastic Leukemia: Differential DiagnosisIma OhwNo ratings yet
- Fungalarthritis and OsteomyelitisDocument21 pagesFungalarthritis and OsteomyelitisYampold Estheben ChusiNo ratings yet
- Um Casamento ArranjadoDocument1 pageUm Casamento ArranjadoJaqueline SantiagoNo ratings yet
- Principles of Disease Control and PreventionDocument13 pagesPrinciples of Disease Control and PreventionAYO NELSON0% (1)
- 3.5 Testing For Syphilis (VDRL)Document3 pages3.5 Testing For Syphilis (VDRL)BALAJINo ratings yet
- Research ProposalDocument4 pagesResearch ProposalAmar GandhiNo ratings yet
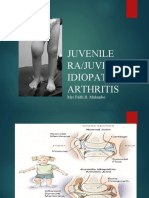
- Juvenile RaDocument27 pagesJuvenile RaEdwardNo ratings yet
- Dengue Duo Engue ns1 Ag Igg Igm SD BiolineDocument4 pagesDengue Duo Engue ns1 Ag Igg Igm SD Biolinersmbgss575No ratings yet
- Vaccination 921007065963Document1 pageVaccination 921007065963ADAMAZAHARINo ratings yet
- Lecture 3 Boils and CarbunclesDocument7 pagesLecture 3 Boils and CarbunclesOsama MalikNo ratings yet
- Correlation Linear Gingival Erythema, Candida Infection and CD4+ Counts in HIV:AIDS Patients at UPIPI RSUD Dr. Soetomo Surabaya, East Java, IndonesiaDocument6 pagesCorrelation Linear Gingival Erythema, Candida Infection and CD4+ Counts in HIV:AIDS Patients at UPIPI RSUD Dr. Soetomo Surabaya, East Java, IndonesiaAchmad Ainul YaqinNo ratings yet
- M-M-R Ii: Swollen GlandsDocument9 pagesM-M-R Ii: Swollen GlandsJoanne Alyssa Hernandez LascanoNo ratings yet
- Aggressive PeriodontitisDocument5 pagesAggressive Periodontitisandreas kevinNo ratings yet
- A13 Microbiology Nursing Course OutlineDocument7 pagesA13 Microbiology Nursing Course OutlinealithebigbossNo ratings yet
- Clinical Presentation On ADDDocument39 pagesClinical Presentation On ADDSREEDEVI T SURESHNo ratings yet
- Blood CellsDocument14 pagesBlood CellsrizaNo ratings yet
- Kode Icd 10Document536 pagesKode Icd 10AkbarisArisNo ratings yet
- Vaccine: Mirriam G.J. Tacken, Franz J. Daus, Femke Feenstra, René G.P. Van Gennip, Piet A. Van RijnDocument7 pagesVaccine: Mirriam G.J. Tacken, Franz J. Daus, Femke Feenstra, René G.P. Van Gennip, Piet A. Van RijnCristina Tommy TomescuNo ratings yet
- Poultry DiseasesDocument13 pagesPoultry DiseasesSajjad LaghariNo ratings yet
- TetanusDocument4 pagesTetanusSneha SnehaNo ratings yet
- 20 Lecture Oral Mucosa Vesiculo-Bullous DiseaseDocument10 pages20 Lecture Oral Mucosa Vesiculo-Bullous Diseaseمازن كريم حمودNo ratings yet