You might also like
- 15 JTH 12782Document12 pages15 JTH 12782Ricardo GomezNo ratings yet
- Sialic Acids On The Surface of Caprine Arthritis-Encephalitis VirusDocument7 pagesSialic Acids On The Surface of Caprine Arthritis-Encephalitis VirusJosé Hiram Sánchez GascaNo ratings yet
- 2022 Certad - CryptosopDocument10 pages2022 Certad - Cryptosopalexandraiuliana.aaNo ratings yet
- The Dynamic Distribution of Duck Tembusu Virus in The Spleen of Infected ShelducksDocument9 pagesThe Dynamic Distribution of Duck Tembusu Virus in The Spleen of Infected ShelducksTrung Kien LeNo ratings yet
- Acute Opisthorchiasis in A Kazakh ManDocument2 pagesAcute Opisthorchiasis in A Kazakh Manhusna aninda farahitaNo ratings yet
- Inhibition of in Vitro Leukocyte Proliferation byDocument7 pagesInhibition of in Vitro Leukocyte Proliferation byIsrael Espinoza HernándezNo ratings yet
- Literature Review On SalmonellaDocument6 pagesLiterature Review On Salmonellac5r9j6zj100% (1)
- Thannesberger 2017Document15 pagesThannesberger 2017Денис КрахоткинNo ratings yet
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus InhibitorsDocument15 pagesThe SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus Inhibitorsfaris daboolNo ratings yet
- Virus Infection Hsp70 Protein Positively Regulates RabiesDocument10 pagesVirus Infection Hsp70 Protein Positively Regulates RabiesAini DollNo ratings yet
- Construction and Application of A Protein Interaction - 2014 - Molecular - CelluDocument14 pagesConstruction and Application of A Protein Interaction - 2014 - Molecular - CelluTanIsmNo ratings yet
- Microbial Pathogenesis: SciencedirectDocument11 pagesMicrobial Pathogenesis: SciencedirectFerdinand Prayogo Cahyo SantosoNo ratings yet
- Ku Rath 2008Document22 pagesKu Rath 2008OanaNo ratings yet
- Pathogens: Herpes Simplex Virus Establishment, Maintenance, and Reactivation: in Vitro Modeling of LatencyDocument14 pagesPathogens: Herpes Simplex Virus Establishment, Maintenance, and Reactivation: in Vitro Modeling of LatencyrehanaNo ratings yet
- Attenuation of Sepsis-Related Immunoparalysis by Continuous Veno-Venous Hemofiltration in Experimental Porcine PancreatitisDocument8 pagesAttenuation of Sepsis-Related Immunoparalysis by Continuous Veno-Venous Hemofiltration in Experimental Porcine PancreatitismanuelantoniomenaNo ratings yet
- Bahan Paper PatoDocument6 pagesBahan Paper PatoKhairul Ihsan Pengusaha MudaNo ratings yet
- 1 or 10 CFU-PMNDocument18 pages1 or 10 CFU-PMNCatherine AtienzaNo ratings yet
- Suraprasit 1Document9 pagesSuraprasit 1Denis ChandraNo ratings yet
- Marroqui2008 Efecto de Inhinidor IPNVDocument7 pagesMarroqui2008 Efecto de Inhinidor IPNVandrea nettle bareaNo ratings yet
- Ann Rev of Virology IPNVDocument30 pagesAnn Rev of Virology IPNVRodolfo VelazcoNo ratings yet
- Apoptosis in Hepatitis C Virus InfectionDocument11 pagesApoptosis in Hepatitis C Virus InfectionHNINNo ratings yet
- Articulo 7 Mig OkDocument11 pagesArticulo 7 Mig Okoscarbio20090% (1)
- Hep 20268Document7 pagesHep 20268ewNo ratings yet
- Jiaa 405Document5 pagesJiaa 405spin_echoNo ratings yet
- Development of The Hybrid Sleeping Beauty-Baculovirus Vector For Sustained Gene Expression and Cancer Therapy 78362006Document9 pagesDevelopment of The Hybrid Sleeping Beauty-Baculovirus Vector For Sustained Gene Expression and Cancer Therapy 78362006Anthony McIvorNo ratings yet
- Pagination VETPAR 109611Document13 pagesPagination VETPAR 109611Bárbara Laurice Araújo VerçosaNo ratings yet
- Combination of Highly Antigenic Nucleoproteins To Inaugurate A Cros - 2021 - HelDocument15 pagesCombination of Highly Antigenic Nucleoproteins To Inaugurate A Cros - 2021 - Helمصطفى الذاريNo ratings yet
- IVDocument12 pagesIVanhtoanly2305No ratings yet
- Aggregating Resistant Hypocoagulability, Hyperfibrinolysis, Phagocytosis, and Neutrophil, Monocyte, and Lymphocyte Binding in Canine Whole BloodDocument15 pagesAggregating Resistant Hypocoagulability, Hyperfibrinolysis, Phagocytosis, and Neutrophil, Monocyte, and Lymphocyte Binding in Canine Whole BloodFabricio CamargoNo ratings yet
- Crimean-Congo Haemorrhagic Fever Virus: MicrobiologyDocument15 pagesCrimean-Congo Haemorrhagic Fever Virus: Microbiologydr sarangNo ratings yet
- 2-Nguyen Et Al.-2014Document7 pages2-Nguyen Et Al.-2014Nguyễn Minh TrangNo ratings yet
- Barrerameja2009 2Document6 pagesBarrerameja2009 2erzebeth bathoryNo ratings yet
- Animal VirologyDocument94 pagesAnimal Virologyabrilama90No ratings yet
- 1 s2.0 S0042682204008438 MainDocument6 pages1 s2.0 S0042682204008438 MainBagus Rahmat BasukiNo ratings yet
- montoya2007Document2 pagesmontoya2007Monica Aleja FernándezNo ratings yet
- Effect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein GeneDocument4 pagesEffect of Antibodies On The Expression of Plasmodium Falciparum Circumsporozoite Protein Genereza gomaNo ratings yet
- Viro 1Document6 pagesViro 1Elsa YuniaNo ratings yet
- Development of An Immuno-Based Colorimetric Assay For White Spot Syndrome VirusDocument6 pagesDevelopment of An Immuno-Based Colorimetric Assay For White Spot Syndrome VirusendorengasNo ratings yet
- Do Protozoa Play A Role in CarcinogenesisDocument6 pagesDo Protozoa Play A Role in CarcinogenesisDaniel GilbertoNo ratings yet
- Within-Host Microevolution of Streptococcus Pneumoniae Is Rapid and Adaptive During Natural Colonisation - 2020 Nature ResearchDocument14 pagesWithin-Host Microevolution of Streptococcus Pneumoniae Is Rapid and Adaptive During Natural Colonisation - 2020 Nature ResearchTony NgNo ratings yet
- RNAi Hepatitis TreatmentDocument12 pagesRNAi Hepatitis TreatmentHironmoy RoyNo ratings yet
- AbstractDocument10 pagesAbstractannisa.ak.1aNo ratings yet
- Stroke in Two Children With Mycoplasma Pneumoniae Infection A Causal or Casual RelationshipDocument15 pagesStroke in Two Children With Mycoplasma Pneumoniae Infection A Causal or Casual Relationshipwawa chenNo ratings yet
- Newcastle Disease Virus ThesisDocument5 pagesNewcastle Disease Virus Thesiskellyratkovicnorman100% (2)
- 基金项目:山东省自然科学基金项目(Y2007D31, Y2007D74);青岛农业大学人才基金项目(630701)。 作者简介:程顺峰,男,1978 年出生,博士,讲师,主要从事水生生物病害学研究。通信地址:266109 山东省青岛市城阳区春阳路青岛农业大学动物科 技学院, 收稿日期:2009-07-02,修回日期:2009-07-24。Document4 pages基金项目:山东省自然科学基金项目(Y2007D31, Y2007D74);青岛农业大学人才基金项目(630701)。 作者简介:程顺峰,男,1978 年出生,博士,讲师,主要从事水生生物病害学研究。通信地址:266109 山东省青岛市城阳区春阳路青岛农业大学动物科 技学院, 收稿日期:2009-07-02,修回日期:2009-07-24。Yunkun RouxNo ratings yet
- 1 s2.0 S1198743X20302895 MainDocument7 pages1 s2.0 S1198743X20302895 MainPhuong NguyenNo ratings yet
- Zebrafish As A Useful Model For Zoonotic Vibrio Parahaemolyticus Pathogenicity in Fish and HumanDocument10 pagesZebrafish As A Useful Model For Zoonotic Vibrio Parahaemolyticus Pathogenicity in Fish and HumanRizka Rahmana PutriNo ratings yet
- Fish and Shell Fish Immunology: Full Length ArticleDocument9 pagesFish and Shell Fish Immunology: Full Length ArticleKirana Rahmada SafitriNo ratings yet
- Sanchez Cordon Et Al 2005 Lymphocyte Apoptosis and Thrombocytopenia in Spleen During Classical Swine Fever Role OfDocument12 pagesSanchez Cordon Et Al 2005 Lymphocyte Apoptosis and Thrombocytopenia in Spleen During Classical Swine Fever Role OfKeto PrehranaNo ratings yet
- Schisto VenezuelaDocument7 pagesSchisto VenezuelaHanna RodagadaNo ratings yet
- Diminished Virulence Rabbit Model of Endocarditis: of Mutant of StaphylococcusDocument8 pagesDiminished Virulence Rabbit Model of Endocarditis: of Mutant of StaphylococcusChrisanti Mau MetaNo ratings yet
- Susceptibility of Channel Catfish Fry To Channel Catfish Virus (CCV) Challenge Increases With AgeDocument9 pagesSusceptibility of Channel Catfish Fry To Channel Catfish Virus (CCV) Challenge Increases With AgeFadhil AusafNo ratings yet
- Lai2015 Patogenitas AHPND Toksin PirDocument9 pagesLai2015 Patogenitas AHPND Toksin PirSuminiNo ratings yet
- A New Nodavirus Is Associated With Covert Mortality Disease of ShrimpDocument10 pagesA New Nodavirus Is Associated With Covert Mortality Disease of ShrimpmiftahuddinmadjidNo ratings yet
- Title: Instructions For UseDocument20 pagesTitle: Instructions For UseAnnisa YohanesNo ratings yet
- Poggio 2015Document10 pagesPoggio 2015Fede JazzNo ratings yet
- Alterations in Macrophage-Produced CytokinesDocument8 pagesAlterations in Macrophage-Produced CytokinessanielNo ratings yet
- What Is Your Diagnosis? Blood Smear From A Cat: Mary Leissinger, Stuart Walton, Jon Fletcher, Stephen GauntDocument2 pagesWhat Is Your Diagnosis? Blood Smear From A Cat: Mary Leissinger, Stuart Walton, Jon Fletcher, Stephen GauntGharbi KamiliaNo ratings yet
- Imunohistokimia Streptavidin Biotin Pada Ikan Nila (Oreochromis Niloticus)Document7 pagesImunohistokimia Streptavidin Biotin Pada Ikan Nila (Oreochromis Niloticus)tiaraNo ratings yet
- Physical Virology: Virus Structure and MechanicsFrom EverandPhysical Virology: Virus Structure and MechanicsUrs F. GreberNo ratings yet
- 2010 Review ReadinessDocument75 pages2010 Review ReadinessGenesis Niogan100% (2)
- APC Pharmacy Intern Written Exam SampleDocument62 pagesAPC Pharmacy Intern Written Exam SampleDaisy Lu81% (16)
- 2013 PATH3206 Course Manual PDFDocument71 pages2013 PATH3206 Course Manual PDFTaonga ZuluNo ratings yet
- International Journal of Pharmaceutical Science Invention (IJPSI)Document6 pagesInternational Journal of Pharmaceutical Science Invention (IJPSI)inventionjournalsNo ratings yet
- Pediatric CAP GuidelineDocument1 pagePediatric CAP GuidelineJohn Vincent Dy OcampoNo ratings yet
- Hip DislocationDocument39 pagesHip DislocationJordan Garcia Aguilar0% (1)
- Janisthaaivfcenter Wordpress Com 2023-11-08 How To Increase The Efficiency of Ivf by Lifestyle ChangesDocument2 pagesJanisthaaivfcenter Wordpress Com 2023-11-08 How To Increase The Efficiency of Ivf by Lifestyle ChangesjanisthaapharmaNo ratings yet
- Meda QantasDocument12 pagesMeda QantasAviasiNo ratings yet
- Pharmacist Interview QuestionsDocument5 pagesPharmacist Interview Questionsmohdshadan87399No ratings yet
- Oppenheim J.J., Feldmann M. - Introduction To The Role of Cytokines in Innate Host Defense and Adaptive Immunity (2000) PDFDocument18 pagesOppenheim J.J., Feldmann M. - Introduction To The Role of Cytokines in Innate Host Defense and Adaptive Immunity (2000) PDFHesbon MomanyiNo ratings yet
- TEACHING PLAN FOR Varicose VeinsDocument5 pagesTEACHING PLAN FOR Varicose Veinscertified_maharot_ako2828No ratings yet
- Recognized By:: MNR Homoeopathic Medical College & Hospital, SangareddyDocument21 pagesRecognized By:: MNR Homoeopathic Medical College & Hospital, SangareddyJahnaveeNo ratings yet
- Rapid HIV Test Detects Antibodies in BloodDocument4 pagesRapid HIV Test Detects Antibodies in Bloodkbl27No ratings yet
- DNC 2Document6 pagesDNC 2Maria VisitacionNo ratings yet
- Medical Data Base Bahrain 2018Document122 pagesMedical Data Base Bahrain 2018Shibu KavullathilNo ratings yet
- Three Treasures Correspondences BrochureDocument4 pagesThree Treasures Correspondences Brochure292293709No ratings yet
- Large Bowel Obstruction GuideDocument13 pagesLarge Bowel Obstruction Guideurmila prajapatiNo ratings yet
- DRUG PRESENTATION On Emergency and CPR DrugsDocument39 pagesDRUG PRESENTATION On Emergency and CPR DrugsShibi BennyNo ratings yet
- OSCDDocument331 pagesOSCDmanselsap100% (1)
- Ashhad's Step 2 CK UW Notes PDFDocument166 pagesAshhad's Step 2 CK UW Notes PDFabNo ratings yet
- 2020 Vision Without Glasses PDFDocument340 pages2020 Vision Without Glasses PDFJacklynlim Lkc100% (3)
- Pediatric Nursing Course OutlineDocument6 pagesPediatric Nursing Course OutlineDaryl Adrian RecaidoNo ratings yet
- WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues PDFDocument423 pagesWHO Classification of Tumours of Haematopoietic and Lymphoid Tissues PDFAlejandra Arely López NegreteNo ratings yet
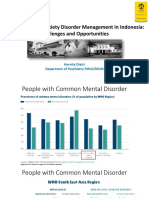
- Hervita - Depression and Anxiety Disorder Management in IndonesiaDocument30 pagesHervita - Depression and Anxiety Disorder Management in IndonesiaPrissilma TaniaNo ratings yet
- Koenig Pastoral Counseling With The AgedDocument11 pagesKoenig Pastoral Counseling With The AgedDavid Danao Franco MendezNo ratings yet
- Healthcare EnvironmentDocument24 pagesHealthcare EnvironmentJOHN VINCENT BJORN GRAGEDA100% (1)
- Hepatitis Case StudyDocument4 pagesHepatitis Case StudyTasha MageeNo ratings yet
- Drug of Choice and DiseasesDocument2 pagesDrug of Choice and DiseasesJamesNo ratings yet
- Yasmeen FatimaDocument11 pagesYasmeen FatimaAdil49No ratings yet
- S-48B Ultrasonic Liposuction Equipment User ManualDocument11 pagesS-48B Ultrasonic Liposuction Equipment User ManualmoparlatinoNo ratings yet