You might also like
- Pediatric Hand: Okta Permata Putri 71 2017 074Document34 pagesPediatric Hand: Okta Permata Putri 71 2017 074Okta PermataNo ratings yet
- EmbryologyDocument26 pagesEmbryologyMeer BabanNo ratings yet
- 10 1016@j Hansur 2018 03 007Document10 pages10 1016@j Hansur 2018 03 007Marcelo CortesNo ratings yet
- Developmental Anomalies of MSK System, 19-20 PDFDocument40 pagesDevelopmental Anomalies of MSK System, 19-20 PDFahmedNo ratings yet
- Lecture 3: Musculoskeletal Development Slide 1Document9 pagesLecture 3: Musculoskeletal Development Slide 1Dionix CruzNo ratings yet
- Pathogenesis of HoloprosencephalyDocument11 pagesPathogenesis of HoloprosencephalyserubimNo ratings yet
- Obs 03Document33 pagesObs 03Alejo VasquezNo ratings yet
- Etyoloji ReviewDocument7 pagesEtyoloji ReviewBerk DurmazNo ratings yet
- Appendicular & LimbDocument29 pagesAppendicular & LimbPeter PrestonNo ratings yet
- BJSTR MS Id 0022673Document16 pagesBJSTR MS Id 0022673Anushka RaipureNo ratings yet
- Diseases of Middle Ear CavityDocument235 pagesDiseases of Middle Ear CavityDr. T. BalasubramanianNo ratings yet
- Daluiski 2001Document15 pagesDaluiski 2001Marcelo CortesNo ratings yet
- Elective EntropyDocument21 pagesElective EntropyDavidroy IRITSNo ratings yet
- M06 Axisspecificationindrosophila Quad 1Document18 pagesM06 Axisspecificationindrosophila Quad 1Mahendra KatreNo ratings yet
- First and Second Branchial Arch Syndromes: Multimodality ApproachDocument38 pagesFirst and Second Branchial Arch Syndromes: Multimodality ApproachDoina Ababii UrsanNo ratings yet
- Supernumerary Digits of The Hand: Eplasty January 2016Document7 pagesSupernumerary Digits of The Hand: Eplasty January 2016Wara Samsarga GedeNo ratings yet
- Congenital ScoliosisDocument41 pagesCongenital ScoliosisnadiaryNo ratings yet
- HugesssDocument4 pagesHugessssmith patelNo ratings yet
- Developmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellDocument3 pagesDevelopmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellAroa ChansNo ratings yet
- 11 Axis FormationDocument61 pages11 Axis Formationapi-3800038100% (1)
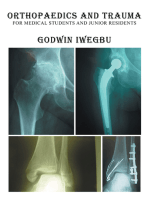
- Orthopaedics and Trauma for Medical Students and Junior ResidentsFrom EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsRating: 5 out of 5 stars5/5 (3)
- Chapter CD: Molecular Control of The Development of Some OrgansDocument16 pagesChapter CD: Molecular Control of The Development of Some OrgansZoher PainterNo ratings yet
- The Developmental Field Concept Genetics in Clinical: Helena, MontDocument5 pagesThe Developmental Field Concept Genetics in Clinical: Helena, MontRi Mar LGNo ratings yet
- Hip Disorders in Children: Postgraduate Orthopaedics SeriesFrom EverandHip Disorders in Children: Postgraduate Orthopaedics SeriesRating: 3 out of 5 stars3/5 (2)
- Radiographic Assessment of Congenital Malformations of The Upper ExtremityDocument17 pagesRadiographic Assessment of Congenital Malformations of The Upper ExtremityZoe GraceNo ratings yet
- Anomalies of Skeletal System-1Document44 pagesAnomalies of Skeletal System-1Meena KoushalNo ratings yet
- Chapter 7: Hand, Fingers and Thumb AnatomyDocument20 pagesChapter 7: Hand, Fingers and Thumb AnatomyRustashNo ratings yet
- Normal Sonogram - SpineDocument4 pagesNormal Sonogram - SpineDuchung TranNo ratings yet
- Lobulated Spleen With A Fissure On Diaphragmatic Surface: RD RDDocument7 pagesLobulated Spleen With A Fissure On Diaphragmatic Surface: RD RDShantu ShirurmathNo ratings yet
- Dental Distress Syndrome QuantifiedDocument29 pagesDental Distress Syndrome QuantifiedNenad Petrövic100% (1)
- Soleus - PhysiopediaDocument6 pagesSoleus - PhysiopediaVahid AtaeiNo ratings yet
- Embryology HandDocument32 pagesEmbryology HandProfesseur Christian DumontierNo ratings yet
- Brachial Plexus InjuryDocument10 pagesBrachial Plexus InjurySanditya FadliNo ratings yet
- Principles and Management of Acute Orthopaedic Trauma: Third EditionFrom EverandPrinciples and Management of Acute Orthopaedic Trauma: Third EditionNo ratings yet
- Yothe Growth in Width of The Facial SkeletonDocument6 pagesYothe Growth in Width of The Facial Skeletonalejac1810No ratings yet
- 2012 Limb DevlopmentDocument50 pages2012 Limb Devlopmentkritimahajan1989No ratings yet
- Anatomy Summary BasicDocument54 pagesAnatomy Summary BasicDungani AllanNo ratings yet
- FMS II DEv of LimbDocument47 pagesFMS II DEv of LimbJoshua ObrienNo ratings yet
- Developmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw SkeletonDocument11 pagesDevelopmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw Skeletonhennysusanto18No ratings yet
- Arthrogryposis (Multiple Congenital Contractures) - Diagnostic Approach To Etiology, Classification, Genetics, and General PrinciplesDocument9 pagesArthrogryposis (Multiple Congenital Contractures) - Diagnostic Approach To Etiology, Classification, Genetics, and General PrinciplesKiara VásquezNo ratings yet
- Bing MentahanDocument15 pagesBing Mentahansecondfery2021No ratings yet
- Epigenetics Assignment - SS24 - Reduced and RevisedDocument14 pagesEpigenetics Assignment - SS24 - Reduced and Revisedbazabrain44No ratings yet
- Genetics of Axis Specification in DrosophilaDocument3 pagesGenetics of Axis Specification in DrosophilaJohnkevin NocheNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The Foregutproject-247758No ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- MRI of the Upper Extremity: Elbow, Wrist, and HandFrom EverandMRI of the Upper Extremity: Elbow, Wrist, and HandBethany U. CasagrandaNo ratings yet
- 3rd Week of DevelopmentDocument20 pages3rd Week of DevelopmentSidra AslamNo ratings yet
- Tratamiento Quirurgico de La SindactiliaDocument13 pagesTratamiento Quirurgico de La SindactiliaJose Alejandro Cobas PazNo ratings yet
- Malformed Vertebrae: A Clinical and Imaging Review: EmbryologyDocument13 pagesMalformed Vertebrae: A Clinical and Imaging Review: EmbryologyYovanka Naryai ManuhutuNo ratings yet
- แกะเทป Developmental of Musculoskeletal SystemDocument50 pagesแกะเทป Developmental of Musculoskeletal Systemjulesarojinee100% (1)
- AxolotlsDocument15 pagesAxolotlsagathapavlovaNo ratings yet
- Development of LimbsDocument29 pagesDevelopment of LimbsRoshan Kumar PanditNo ratings yet
- Epos Hip UltrasoundDocument20 pagesEpos Hip UltrasoundBogdan Popescu100% (1)
- Cleft Lip and Palate Management: A Comprehensive AtlasFrom EverandCleft Lip and Palate Management: A Comprehensive AtlasRicardo D. BennunNo ratings yet
- 10.1007@s00247 019 04512 3 PDFDocument12 pages10.1007@s00247 019 04512 3 PDFpablo gonzalezNo ratings yet
- Acb 46 149Document8 pagesAcb 46 149soylahijadeunvampiroNo ratings yet
- Morphogenetic Mechanisms1Document36 pagesMorphogenetic Mechanisms1Lily VaughNo ratings yet
- Anatomical Variation in The Formation of Median Nerve - A Case ReportDocument3 pagesAnatomical Variation in The Formation of Median Nerve - A Case ReportIOSR Journal of PharmacyNo ratings yet
- DM Infectious DiseasesDocument20 pagesDM Infectious DiseasesVasishta NadellaNo ratings yet
- The Strawberry Tongue: What, How and WhereDocument6 pagesThe Strawberry Tongue: What, How and WhereVasishta NadellaNo ratings yet
- Mobile Medical UnitsDocument100 pagesMobile Medical UnitsVasishta NadellaNo ratings yet
- Vaccines & DiluentsDocument1 pageVaccines & DiluentsVasishta NadellaNo ratings yet
- Baterial & Fungal Corneal UlcersDocument28 pagesBaterial & Fungal Corneal UlcersVasishta NadellaNo ratings yet
- Emesis in The NewbornDocument34 pagesEmesis in The NewbornVasishta NadellaNo ratings yet
- Neuromuscular Disorders: Jean-Claude KaplanDocument22 pagesNeuromuscular Disorders: Jean-Claude KaplanVasishta NadellaNo ratings yet
- IGH (Indian Guidelines On Hypertension) - 2019 GUIDELINESDocument15 pagesIGH (Indian Guidelines On Hypertension) - 2019 GUIDELINESVasishta NadellaNo ratings yet
- Cold Chain: Procurement GuidelinesDocument12 pagesCold Chain: Procurement GuidelinesVasishta NadellaNo ratings yet
- Who Can Access Research4Life?: Group A Countries, Areas and Territories - Free AccessDocument2 pagesWho Can Access Research4Life?: Group A Countries, Areas and Territories - Free AccessVasishta NadellaNo ratings yet
- Abnormal Head Position PDFDocument2 pagesAbnormal Head Position PDFVasishta NadellaNo ratings yet
- Mechanism of Wenchebach Type AV BlockDocument11 pagesMechanism of Wenchebach Type AV BlockVasishta NadellaNo ratings yet
- Brucella SpeciesDocument15 pagesBrucella SpeciesVasishta NadellaNo ratings yet
- Flow Cytometry Basics GuideDocument72 pagesFlow Cytometry Basics GuideVasishta NadellaNo ratings yet
- Paradoxical Head Tilt in Unilateral Traumatic Superior Oblique PalsyDocument3 pagesParadoxical Head Tilt in Unilateral Traumatic Superior Oblique PalsyVasishta NadellaNo ratings yet
- Psammoma BodyDocument8 pagesPsammoma BodyVasishta NadellaNo ratings yet
- Pituitary Gland Pathology: Best Practice No 172Document8 pagesPituitary Gland Pathology: Best Practice No 172Vasishta NadellaNo ratings yet
- Sinus Node DysfunctionDocument11 pagesSinus Node DysfunctionVasishta Nadella100% (1)
- Hunner Lesion (Formerly Known As Hunner'S Ulcer)Document4 pagesHunner Lesion (Formerly Known As Hunner'S Ulcer)Vasishta NadellaNo ratings yet
- Antiepileptic Drug ToleranceDocument3 pagesAntiepileptic Drug ToleranceVasishta NadellaNo ratings yet
- Overview of Established Antiepileptic DrugsDocument13 pagesOverview of Established Antiepileptic DrugsVasishta NadellaNo ratings yet
- Assessment of Blood Loss During Cesarean Section Under General Anesthesia and Epidural Analgesia Using Different MethodsDocument10 pagesAssessment of Blood Loss During Cesarean Section Under General Anesthesia and Epidural Analgesia Using Different MethodsVasishta NadellaNo ratings yet
- MRKH Jurnal SHDocument9 pagesMRKH Jurnal SHSUSANNo ratings yet
- Polyspermy and How To Reduce ItDocument35 pagesPolyspermy and How To Reduce ItbrachyotisNo ratings yet
- City of Malaybalay: San Isidro College Integrated Basic EducationDocument3 pagesCity of Malaybalay: San Isidro College Integrated Basic EducationJessie Cris GumonayNo ratings yet
- Sexual Reproduction LPDocument15 pagesSexual Reproduction LPKATRINA MARIE AmayNo ratings yet
- 1196-Article Text-5855-1-10-20221222Document11 pages1196-Article Text-5855-1-10-20221222vinna munadNo ratings yet
- The First SexDocument399 pagesThe First SexAcenda O Espírito100% (1)
- Biology: Unit: KBI0/4BI0 Paper: 2BDocument20 pagesBiology: Unit: KBI0/4BI0 Paper: 2BMohamedNo ratings yet
- Wiersma, Rinus PDFDocument164 pagesWiersma, Rinus PDFΔημήτρης ΓεωργίουNo ratings yet
- MeiosisDocument13 pagesMeiosistaylorNo ratings yet
- Class 12 Bio Zoology TM Chapter-1 Full GuideDocument7 pagesClass 12 Bio Zoology TM Chapter-1 Full GuidemakeshwaranNo ratings yet
- Artigo Pra Ler 1Document10 pagesArtigo Pra Ler 1Luiz ClaudioNo ratings yet
- Midterms Reviewer Gee11Document2 pagesMidterms Reviewer Gee11HarimasuNo ratings yet
- Maternity Nursing An Introductory Text 11th Edition Leifer Test BankDocument35 pagesMaternity Nursing An Introductory Text 11th Edition Leifer Test Bankmoonseedegoismwjvm695% (21)
- Development of The Vertebrate Forelimb - Power Point (Group 28)Document29 pagesDevelopment of The Vertebrate Forelimb - Power Point (Group 28)mayreaupatriotNo ratings yet
- UrochordatevolutionDocument15 pagesUrochordatevolutionRodrigo CarralNo ratings yet
- The Way of The CellDocument320 pagesThe Way of The Cellhuynhthiensach100% (2)
- Estrous CycleDocument3 pagesEstrous CyclehumanupgradeNo ratings yet
- Simone de Beauvoir - The Second Sex PDFDocument210 pagesSimone de Beauvoir - The Second Sex PDFmaieruchan100% (1)
- Female Reproductive System Lab-2021Document37 pagesFemale Reproductive System Lab-2021Oo OoNo ratings yet
- Development of Face &it's Applied AspectsDocument95 pagesDevelopment of Face &it's Applied AspectsHansa KunduNo ratings yet
- Sexual IntercourseDocument5 pagesSexual IntercourseshitfikapeNo ratings yet
- 18 Hour Chick EmbryoDocument7 pages18 Hour Chick Embryoaa628100% (2)
- Second Periodical Test - For ReDocument4 pagesSecond Periodical Test - For ReJessica Manawes Navor100% (1)
- 2019 Sec 4 Pure Biology-12sDocument571 pages2019 Sec 4 Pure Biology-12sUZAIR MAHBUB BHUYAINNo ratings yet
- Ob Review QuestionsDocument31 pagesOb Review QuestionsRandyNo ratings yet
- Reproduction in Organisms-1Document52 pagesReproduction in Organisms-1Dynemech Anti Vibration TechnologyNo ratings yet
- VOLVOXDocument25 pagesVOLVOXTOM JOSEPHNo ratings yet
- InfertiityDocument36 pagesInfertiityKavya S Mohan100% (1)
- Gestational Trophoblastic DiseaseDocument4 pagesGestational Trophoblastic DiseasePrincess PlateroNo ratings yet
- General Biology 2 IMDocument215 pagesGeneral Biology 2 IMPatrick Verroya100% (1)