You might also like
- O. Tauri: 3-D Ultrastructure Of: Electron Cryotomography of An Entire Eukaryotic CellDocument14 pagesO. Tauri: 3-D Ultrastructure Of: Electron Cryotomography of An Entire Eukaryotic CellMARIA ANGGIE CANTIKA DEWANINo ratings yet
- Hall CyanelaDocument13 pagesHall CyanelaGabyNo ratings yet
- From The Laboratories of The Rockefdler Inai Ute For M e D ResearchDocument39 pagesFrom The Laboratories of The Rockefdler Inai Ute For M e D Researchnorok77560No ratings yet
- Micro e Nanoestrutura CelularDocument4 pagesMicro e Nanoestrutura CelularIsis CôrtesNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- From Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los AngelesDocument15 pagesFrom Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los Angelesarclite86No ratings yet
- Encyclopediabookchapter PDFDocument13 pagesEncyclopediabookchapter PDFJamesNo ratings yet
- Figure 4-9. (Alberts4e) A Cross-Sectional View of A Typical Cell NucleusDocument8 pagesFigure 4-9. (Alberts4e) A Cross-Sectional View of A Typical Cell NucleusLaura RecaldeNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- 16 Staehelin PDFDocument12 pages16 Staehelin PDFAmandaPutriNastitiNo ratings yet
- Phycology: DinophytaDocument76 pagesPhycology: Dinophytamichael yinNo ratings yet
- Chromosomes and Nucleoli of The Axolotl, Ambystoma MexicanumDocument40 pagesChromosomes and Nucleoli of The Axolotl, Ambystoma MexicanumBrayan Anaya LavalleNo ratings yet
- Fine Structure and Morphogenic Movements in The Gastrula of The Treefrog, Hyla RegillaDocument24 pagesFine Structure and Morphogenic Movements in The Gastrula of The Treefrog, Hyla Regillaimanuel kuneNo ratings yet
- Dent 1961Document11 pagesDent 1961Brayden EaglesNo ratings yet
- From The Departnwnl of Anatomy, University of Washington, SeattleDocument17 pagesFrom The Departnwnl of Anatomy, University of Washington, Seattlenorok77560No ratings yet
- Jnnpsyc00297 0079Document8 pagesJnnpsyc00297 0079EccoNo ratings yet
- The Epithelial Basement MembraneDocument9 pagesThe Epithelial Basement MembraneSitipradyta KasimNo ratings yet
- Chloroplast: Jump To Navigationjump To SearchDocument3 pagesChloroplast: Jump To Navigationjump To SearchdumbassNo ratings yet
- J Bbagrm 2009 02 002Document7 pagesJ Bbagrm 2009 02 002Raymundo Lopez NNo ratings yet
- GembiologyDocument10 pagesGembiologyalexymersNo ratings yet
- Chloroplast EvolutionDocument3 pagesChloroplast EvolutionobaytutNo ratings yet
- CHP 2 Structure and Function of CellsDocument29 pagesCHP 2 Structure and Function of CellsFathima NusrathNo ratings yet
- The Inferior Olivary Nucleus of The Rat A Light and Electron Microscopic StudyDocument31 pagesThe Inferior Olivary Nucleus of The Rat A Light and Electron Microscopic StudyRafael GutierrezNo ratings yet
- 04 Prokaryotic Algae ProcholopytesDocument17 pages04 Prokaryotic Algae Procholopytesanne ropNo ratings yet
- ChloroplastDocument35 pagesChloroplastmymobile502No ratings yet
- Archaebacteria (Archaea) and The Origin of The Eukaryotic NucleusDocument8 pagesArchaebacteria (Archaea) and The Origin of The Eukaryotic NucleusDeboraRikaAngelitaNo ratings yet
- Edidin 202003Document6 pagesEdidin 202003david torresNo ratings yet
- Chloroplast: Sterols CyanidiumDocument3 pagesChloroplast: Sterols CyanidiumApriani BinurNo ratings yet
- Anatomy and Physiology of The Protozoa: Chapter TwoDocument31 pagesAnatomy and Physiology of The Protozoa: Chapter TwoHassan AL-HillaliNo ratings yet
- A Chloroplast: ArabidopsisDocument2 pagesA Chloroplast: ArabidopsisRamesh AdhikariNo ratings yet
- Kalinina Protistology 12-1Document9 pagesKalinina Protistology 12-1jesus javier hoyos acuñaNo ratings yet
- Cell Structure and Function Matching KEYDocument3 pagesCell Structure and Function Matching KEYJustin Lin100% (1)
- Photosynthesis & LeavesDocument38 pagesPhotosynthesis & LeavesSanthoshNo ratings yet
- Cellular Structures of Porpbyridium CruentumDocument11 pagesCellular Structures of Porpbyridium CruentumgallondNo ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- Elife 84974 v2Document24 pagesElife 84974 v2Anna McMeekinsNo ratings yet
- Concepts of Membrane StructureFrom EverandConcepts of Membrane StructureRonald AloiaNo ratings yet
- The Ultrastructure of the Animal Cell: International Series in Pure and Applied Biology, Volume 55From EverandThe Ultrastructure of the Animal Cell: International Series in Pure and Applied Biology, Volume 55No ratings yet
- TMP A6 AFDocument9 pagesTMP A6 AFFrontiersNo ratings yet
- Corneal Epithelial Stem Cells: Deficiency and Regulation: # Humana Press 2008Document10 pagesCorneal Epithelial Stem Cells: Deficiency and Regulation: # Humana Press 2008Erick MartinezNo ratings yet
- Cells: H1N3 Publications SDN BHD®Document22 pagesCells: H1N3 Publications SDN BHD®Noor HafizNo ratings yet
- Desarrollo Del CloroplastoDocument21 pagesDesarrollo Del CloroplastoAnderson HernándezNo ratings yet
- Peg-Dextren 1990Document1 pagePeg-Dextren 1990Hass GraceNo ratings yet
- 2019 Klug Et Al AnatomyEvolutionFirstColeoidea Communications BiologyDocument13 pages2019 Klug Et Al AnatomyEvolutionFirstColeoidea Communications BiologyEnrique Bernardez RodriguezNo ratings yet
- Fluid Mosaic ModelDocument6 pagesFluid Mosaic Modelhotlipz_126No ratings yet
- Biologi Molekuler - Lipid BilayerDocument1 pageBiologi Molekuler - Lipid BilayerCinsy PaskalineNo ratings yet
- Raviteja1 PDFDocument6 pagesRaviteja1 PDFMekala LakshmanNo ratings yet
- First Discussion Period : D. Dr. Dr. IDocument9 pagesFirst Discussion Period : D. Dr. Dr. IEusebio ChaconNo ratings yet
- Regulation of Crustacean Molting-A Review and Our PerspectivesDocument8 pagesRegulation of Crustacean Molting-A Review and Our PerspectivesJ HNo ratings yet
- Cell Biology NotesDocument35 pagesCell Biology NotesShreyank TomarNo ratings yet
- NIH Public Access: Photoreceptor Structure and Development: Analyses Using GFP TransgenesDocument12 pagesNIH Public Access: Photoreceptor Structure and Development: Analyses Using GFP TransgenesBps bidan esumNo ratings yet
- An Electron Microscope Study of Thin Sections of Human Skin LPADocument21 pagesAn Electron Microscope Study of Thin Sections of Human Skin LPALucas PolizzeliNo ratings yet
- Vakuola Kontraktil PDFDocument17 pagesVakuola Kontraktil PDFShin Oriflame MalangNo ratings yet
- Fung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHDocument6 pagesFung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHcarciclaroNo ratings yet
- Biology 9th PracticalDocument22 pagesBiology 9th PracticalMansha KhanNo ratings yet
- Taton2011 Article PlectolyngbyaHodgsoniiANovelFiDocument11 pagesTaton2011 Article PlectolyngbyaHodgsoniiANovelFiHugo SantanaNo ratings yet
- Sick and TiredDocument24 pagesSick and TiredVaradrajan MudliyarNo ratings yet
- Laboratory ActivitiesDocument12 pagesLaboratory Activitiesjovanalyn mananganNo ratings yet
- Microbiology Fundamentals A Clinical Approach 1st Edition Cowan Solutions ManualDocument36 pagesMicrobiology Fundamentals A Clinical Approach 1st Edition Cowan Solutions Manualduongnujl33q100% (24)
- Antimicrobial Effects of Airborne Acoustic Ultrasound and Plasma Activated Water From Cold and Thermal Plasma Systems On BiofilmsDocument10 pagesAntimicrobial Effects of Airborne Acoustic Ultrasound and Plasma Activated Water From Cold and Thermal Plasma Systems On BiofilmsJulio MoralesNo ratings yet
- Morphology and Function of Malpighian Tubules and Associated Structures in The Cockroach, Periplaneta AmericanaDocument42 pagesMorphology and Function of Malpighian Tubules and Associated Structures in The Cockroach, Periplaneta AmericanaManojit ChatterjeeNo ratings yet
- Comparison of The Cytotoxicity of Formocresol, Formaldehyde, Cresol, and Glutaraldehyde Using Human Pulp Fibroblast CulturesDocument6 pagesComparison of The Cytotoxicity of Formocresol, Formaldehyde, Cresol, and Glutaraldehyde Using Human Pulp Fibroblast Culturesvj297No ratings yet
- Special StainsDocument10 pagesSpecial Stainsjgpd2000100% (1)
- Introduction To StainingDocument15 pagesIntroduction To Stainingitzel rendonNo ratings yet
- Fixing, Staining and MicrosDocument2 pagesFixing, Staining and Microshajar ahmadNo ratings yet
- US Army Medical Course MD0856-200 - BacteriologyDocument267 pagesUS Army Medical Course MD0856-200 - BacteriologyGeorges100% (2)
- Textbook of Oral Pathology-Anil G. Ghom, S. Mhaske (2013)Document1,050 pagesTextbook of Oral Pathology-Anil G. Ghom, S. Mhaske (2013)Khánh Vũ100% (2)
- MIC159 Microbial WorldDocument36 pagesMIC159 Microbial WorldDegee GonzalesNo ratings yet
- FYBSC MicrobiologyDocument12 pagesFYBSC MicrobiologyHansa Boricha100% (1)
- Final Syllabi - Microbiology 10062019Document57 pagesFinal Syllabi - Microbiology 10062019JharkhandNo ratings yet
- Dedication I Want To Dedicate This SIWES Report To God Almighty The Owner of My Life For His Love andDocument30 pagesDedication I Want To Dedicate This SIWES Report To God Almighty The Owner of My Life For His Love andAkinrinade Abisola OmopelolaNo ratings yet
- (MT 57) para - MidtermsDocument56 pages(MT 57) para - MidtermsLiana JeonNo ratings yet
- Microbiology and ParasitologyDocument3 pagesMicrobiology and ParasitologyGeorgia LanuzoNo ratings yet
- Bapat Et Al. - 2006 - Quantification of Metabolically Active Biomass Using Methylene Blue Dye Reduction Test (MBRT) Measurement of CFU IDocument10 pagesBapat Et Al. - 2006 - Quantification of Metabolically Active Biomass Using Methylene Blue Dye Reduction Test (MBRT) Measurement of CFU IJosé Matías ZapiolaNo ratings yet
- Piis0272638618311028 PDFDocument12 pagesPiis0272638618311028 PDFRobin GonzalezNo ratings yet
- ToplabDocument16 pagesToplabPaunescu Ioan CornelNo ratings yet
- Biophotons As Neural Communication Signals PDFDocument8 pagesBiophotons As Neural Communication Signals PDFweb3351No ratings yet
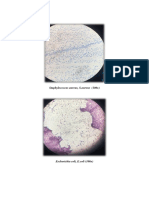
- Result:: Staphylococcus Aureus, S.aureus (100x)Document3 pagesResult:: Staphylococcus Aureus, S.aureus (100x)Ayuni Nadrah Bt KamarujamanNo ratings yet
- PMLS Chapter 2 - Defining The Practice of He Medical Technology - Clinical Laboratory Science ProfessionDocument8 pagesPMLS Chapter 2 - Defining The Practice of He Medical Technology - Clinical Laboratory Science ProfessionRamela Nicole AldaveNo ratings yet
- Rajan Et Al., 2013, Galectin-1 Gadus MorhuaDocument11 pagesRajan Et Al., 2013, Galectin-1 Gadus MorhuaLulu SetiyabudiNo ratings yet
- Histological Changes Associated With Consumption of TanjorensisDocument10 pagesHistological Changes Associated With Consumption of TanjorensisNya Joshua AsuquoNo ratings yet
- General Zoology Laboratory Exercise 2Document5 pagesGeneral Zoology Laboratory Exercise 2Wallen LagradaNo ratings yet
- Gram's Stain ReagentDocument3 pagesGram's Stain Reagentsubhaga dubeyNo ratings yet
- Tuberculosis Diagnosis-HiMediaDocument12 pagesTuberculosis Diagnosis-HiMediaflorengrace.fg98No ratings yet
- Review of Medical Microbiology and Immunology 17th Edition (Medicalstudyzone - Com)Document1,016 pagesReview of Medical Microbiology and Immunology 17th Edition (Medicalstudyzone - Com)FaizNo ratings yet
- Lesson 08Document9 pagesLesson 08asyafujisaki100% (2)