You might also like
- Corneal Epithelial Wound Healing: PerspectiveDocument8 pagesCorneal Epithelial Wound Healing: PerspectiveHanaNo ratings yet
- Structure and Function of Membrane Proteins: Proceedings of the International Symposium on Structure and Function of Membrane Proteins Held in Selva Di Fasano (Italy), May 23-26, 1983From EverandStructure and Function of Membrane Proteins: Proceedings of the International Symposium on Structure and Function of Membrane Proteins Held in Selva Di Fasano (Italy), May 23-26, 1983E. QuagliarielloNo ratings yet
- Corneal LayerDocument9 pagesCorneal LayerAriNo ratings yet
- Ahmad 2007Document11 pagesAhmad 2007Erick MartinezNo ratings yet
- Keratinization of The Oral EpitheliumDocument8 pagesKeratinization of The Oral EpitheliumZachary DuongNo ratings yet
- Membrane Models: Model That Is Accepted TodayDocument4 pagesMembrane Models: Model That Is Accepted TodayWaqas KhanNo ratings yet
- A6. Karring T, Lang N, Löe H. The Role of Gingival Connective Tissue in Determining Epithelial Differentiation. J Perio Res 1975Document12 pagesA6. Karring T, Lang N, Löe H. The Role of Gingival Connective Tissue in Determining Epithelial Differentiation. J Perio Res 1975Daniel CubasNo ratings yet
- Electron Microscopy Study Human Skin Cell Layers Footpad EpidermisDocument21 pagesElectron Microscopy Study Human Skin Cell Layers Footpad EpidermisLucas PolizzeliNo ratings yet
- 1970 - Kollar - DP Embrionic FormationDocument5 pages1970 - Kollar - DP Embrionic FormationalexantonjazzNo ratings yet
- A4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Document16 pagesA4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Daniel CubasNo ratings yet
- The glomerular basement membrane: not gone, just forgottenDocument4 pagesThe glomerular basement membrane: not gone, just forgottenalma ramirezNo ratings yet
- Karring 1975Document12 pagesKarring 1975Juan Pablo FloresNo ratings yet
- Epidermólisis BullosaDocument25 pagesEpidermólisis BullosakYYBNo ratings yet
- Gorter and Grendel's Membrane Theory (1920)Document4 pagesGorter and Grendel's Membrane Theory (1920)HARESHNo ratings yet
- Even Tzur2010Document12 pagesEven Tzur2010Rose Sky SiriusNo ratings yet
- The Behavior of Fibroblasts from the Developing Avian CorneaDocument19 pagesThe Behavior of Fibroblasts from the Developing Avian CorneaBastion ReinhartNo ratings yet
- Characterization of A Biomimetic Polymeric Lipid Bilayer by Phase Sensitive Neutron ReflectometryDocument22 pagesCharacterization of A Biomimetic Polymeric Lipid Bilayer by Phase Sensitive Neutron Reflectometrynyc100% (1)
- Course: PG-Pathshala Paper-12: Membrane Biophysics Content Writer: Dr. Jitendraa Vashistt, Jaypee University of Information & Technology, (H.P)Document8 pagesCourse: PG-Pathshala Paper-12: Membrane Biophysics Content Writer: Dr. Jitendraa Vashistt, Jaypee University of Information & Technology, (H.P)Tejinder SinghNo ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- Functional Architecture of BiomembranesDocument5 pagesFunctional Architecture of BiomembranesAuraAuraNo ratings yet
- Pi Is 0006349518321933Document1 pagePi Is 0006349518321933ASP ValenciaNo ratings yet
- Electron Microscopic Features of TheDocument8 pagesElectron Microscopic Features of TheDr. DeeptiNo ratings yet
- Cell and Molecular Biology Concepts and Experiments 7th Edition Karp Solutions Manual 1Document50 pagesCell and Molecular Biology Concepts and Experiments 7th Edition Karp Solutions Manual 1lauren100% (28)
- Ultrastructure of The Skin Melanophores and Iridophores inDocument4 pagesUltrastructure of The Skin Melanophores and Iridophores inTiberiu MarianNo ratings yet
- Functie Si Structura CorneeDocument17 pagesFunctie Si Structura CorneeMaria CorbuNo ratings yet
- Plasma Membrane Structure and FunctionsDocument12 pagesPlasma Membrane Structure and FunctionsThrupthiNo ratings yet
- Technological Institute of The PhilippinesDocument8 pagesTechnological Institute of The PhilippinesJohn Dolph FacundoNo ratings yet
- Cell Modification FunctionsDocument24 pagesCell Modification FunctionsAmado Sarangay Banawa Jr.No ratings yet
- Corneal Epithelial Stem Cells: Deficiency and Regulation: # Humana Press 2008Document10 pagesCorneal Epithelial Stem Cells: Deficiency and Regulation: # Humana Press 2008Erick MartinezNo ratings yet
- Anatomy, Physiology and Biochemistry of Cornea: Presenter-Yordanos (R1) Moderator-Dr YaredDocument106 pagesAnatomy, Physiology and Biochemistry of Cornea: Presenter-Yordanos (R1) Moderator-Dr YaredAmare GetnetNo ratings yet
- Synergy Between Genetic and Tissue Engineering: Creating An Artificial Salivary GlandDocument6 pagesSynergy Between Genetic and Tissue Engineering: Creating An Artificial Salivary Glandammarstein89No ratings yet
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Jean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsDocument12 pagesJean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsHutsDMNo ratings yet
- From Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los AngelesDocument15 pagesFrom Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los Angelesarclite86No ratings yet
- The Anatomy of The LimbusDocument8 pagesThe Anatomy of The LimbusBukaNo ratings yet
- A M I E: Dvanced Ethods N LectrophysiologyDocument43 pagesA M I E: Dvanced Ethods N LectrophysiologyArtur LiberatoNo ratings yet
- Epithelium and Connective Tissue Lecture SummaryDocument4 pagesEpithelium and Connective Tissue Lecture SummaryDan ChoiNo ratings yet
- Cell Anatomy OrganellesDocument7 pagesCell Anatomy OrganellesGoogle BackupNo ratings yet
- Fucus VesiculosusDocument15 pagesFucus VesiculosusRistaNo ratings yet
- Structure of Eukaryotic Cells Drawing Eukaryotic CellsDocument4 pagesStructure of Eukaryotic Cells Drawing Eukaryotic CellsAldo LoyolaNo ratings yet
- Ijos 201346Document9 pagesIjos 201346Arnasary KaharNo ratings yet
- Membrane Models: Gorter and Grendel's Membrane Theory (1920)Document5 pagesMembrane Models: Gorter and Grendel's Membrane Theory (1920)Hafiz AhmadNo ratings yet
- Cell StructureDocument6 pagesCell StructureL.y. Chong100% (1)
- Nucleo Kim and Wirtz Biomaterials 2015Document12 pagesNucleo Kim and Wirtz Biomaterials 2015miriamNo ratings yet
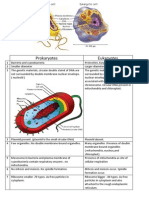
- The Physiology of Prokaryotic Cell: Understanding the Smallest Form of LifeDocument22 pagesThe Physiology of Prokaryotic Cell: Understanding the Smallest Form of LifeAnnysaNurmalaSariNo ratings yet
- Morphological Immunochemical: HypertrophicDocument9 pagesMorphological Immunochemical: HypertrophicRarasati SrimurniNo ratings yet
- Keratinization - Dr. PriyaDocument37 pagesKeratinization - Dr. PriyaDr. Priya Patel75% (4)
- Cell MembraneDocument6 pagesCell MembraneÖmer KhanNo ratings yet
- Dentogingival Junction AnatomyDocument7 pagesDentogingival Junction AnatomyDrKrishna Das100% (1)
- Helium Ion Microscopy - BacteriumDocument8 pagesHelium Ion Microscopy - BacteriumElis SoaresNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture Outlinehaha_le12No ratings yet
- Assignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3ADocument9 pagesAssignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3Ajane austin lynn rebancosNo ratings yet
- Nanofiber-Reinforced Decellularized Amniotic Membrane Improves Limbal Stem Cell Transplantation in A Rabbit Model of Corneal Epithelial DefectDocument11 pagesNanofiber-Reinforced Decellularized Amniotic Membrane Improves Limbal Stem Cell Transplantation in A Rabbit Model of Corneal Epithelial DefectselcanekicierNo ratings yet
- Shoukath2017 Artigo Vasos LinfáticosDocument10 pagesShoukath2017 Artigo Vasos LinfáticosGustavo MunizNo ratings yet
- Cell MembraneDocument23 pagesCell MembraneDoctora NourhanNo ratings yet
- Understandings, Applications and Skills (This Is What You Maybe Assessed On)Document11 pagesUnderstandings, Applications and Skills (This Is What You Maybe Assessed On)yara hazemNo ratings yet
- 2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisDocument16 pages2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisskbucherNo ratings yet
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Chapter 7 Lecture: Cell BiologyDocument86 pagesChapter 7 Lecture: Cell BiologyQueenie Anne Barroga AspirasNo ratings yet
- General Anaesthesia: A Brief History and Guide to Its Mechanism, Stages and ApplicationsDocument19 pagesGeneral Anaesthesia: A Brief History and Guide to Its Mechanism, Stages and ApplicationsSunetraNo ratings yet
- LAS Q3-Health8-Week-6-8Document8 pagesLAS Q3-Health8-Week-6-8novem salonoyNo ratings yet
- Unit 2 - Develop and Implement The Health and Safety Policy Workbook RGDocument11 pagesUnit 2 - Develop and Implement The Health and Safety Policy Workbook RGAshraf EL WardajiNo ratings yet
- Summary Writing: Theory & Practice (2020) : 1 - PageDocument6 pagesSummary Writing: Theory & Practice (2020) : 1 - PageChin Thục HiềnNo ratings yet
- Plan of ActivitiesDocument3 pagesPlan of ActivitiesLiiza G-GsprNo ratings yet
- Main modifications to Phoresis softwareDocument70 pagesMain modifications to Phoresis softwareVu AnhNo ratings yet
- Reviewer in QUIZ 1 2 AND 3 5Document3 pagesReviewer in QUIZ 1 2 AND 3 5John Eldrin DuyagNo ratings yet
- Plastic Surgery ConferenceDocument1 pagePlastic Surgery Conferenceretno widyastutiNo ratings yet
- Evaluation and Comparison of The New Mindray BC-6200 Hematology Analyzer With ADVIA 2120iDocument8 pagesEvaluation and Comparison of The New Mindray BC-6200 Hematology Analyzer With ADVIA 2120iGABRIELA HERRERANo ratings yet
- NCE - Pre - Inter - Module 2 (10) - Take CareDocument14 pagesNCE - Pre - Inter - Module 2 (10) - Take CarePhạm Quang ThiênNo ratings yet
- Prueba 2Document188 pagesPrueba 2David LiNo ratings yet
- EPI Vaccines: - Inactivated (Killed) Microorganisms - Attentuated Microorganisms Fragments From Microorganisms ToxoidsDocument3 pagesEPI Vaccines: - Inactivated (Killed) Microorganisms - Attentuated Microorganisms Fragments From Microorganisms ToxoidsIrene Grace BalcuevaNo ratings yet
- God, Buber, GestaltDocument15 pagesGod, Buber, GestaltAriel Pollak100% (1)
- IEP-Anikka Rebecca Alarcon (DepEd)Document6 pagesIEP-Anikka Rebecca Alarcon (DepEd)may may abelleraNo ratings yet
- CounterclaimsDocument3 pagesCounterclaimsRuthie PatootieNo ratings yet
- Romanian Firefighters NR 4 - 2022Document52 pagesRomanian Firefighters NR 4 - 2022Forum PompieriiNo ratings yet
- Student Sex Work Toolkit FINAL December 2020Document11 pagesStudent Sex Work Toolkit FINAL December 2020due ruoteNo ratings yet
- ABSTRACT - BOOK - 10th IcgsDocument135 pagesABSTRACT - BOOK - 10th Icgsd_kourkoutasNo ratings yet
- Partograph: Source: Basic Emergency Obstetric Care Dr. Jose Fabella Memorial Hospital, & Who By: Araceli F. SuratDocument33 pagesPartograph: Source: Basic Emergency Obstetric Care Dr. Jose Fabella Memorial Hospital, & Who By: Araceli F. SuratRaquel MonsalveNo ratings yet
- Science & Technology WritingDocument92 pagesScience & Technology Writingrandy alvaroNo ratings yet
- The World of Work CrosswordDocument1 pageThe World of Work CrosswordNikolay SpassovNo ratings yet
- Strategy Implementation in Organizations A ConceptDocument14 pagesStrategy Implementation in Organizations A ConceptGemechis LemaNo ratings yet
- SDS-Dolphin Screen WashDocument10 pagesSDS-Dolphin Screen WashLiz CNo ratings yet
- Acrylic Partial DenturesDocument19 pagesAcrylic Partial Denturesahmad955mls100% (1)
- KITCHEN VIPER G I Joe RPGDocument1 pageKITCHEN VIPER G I Joe RPGwoofNo ratings yet
- Active Listening SkillsDocument6 pagesActive Listening SkillssrhmdNo ratings yet
- PCN Certification Services GuideDocument3 pagesPCN Certification Services Guidenarutothunderjet216No ratings yet
- Keeping Yourself Company: Sahil Patel MAY2020Document18 pagesKeeping Yourself Company: Sahil Patel MAY2020Sahil PatelNo ratings yet
- Musclebear, J, R - Eat Clen Tren Hard - The Guide For Steroid Use in Powerlifting (2017)Document75 pagesMusclebear, J, R - Eat Clen Tren Hard - The Guide For Steroid Use in Powerlifting (2017)macajir6980% (2)
- Mental Health AwarenessDocument2 pagesMental Health Awarenessfarhatulhanimzulkifl100% (2)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)