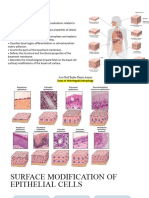
You might also like
- Oral Mucosa: (Ten Cates)Document14 pagesOral Mucosa: (Ten Cates)Vanshika SethiNo ratings yet
- Harrys CosmetologyDocument80 pagesHarrys CosmetologyNidhi100% (3)
- Oral Mucous MembraneDocument71 pagesOral Mucous MembraneHarleen GrewalNo ratings yet
- Keratinization - Dr. PriyaDocument37 pagesKeratinization - Dr. PriyaDr. Priya Patel75% (4)
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Corneal Epithelial Wound Healing: PerspectiveDocument8 pagesCorneal Epithelial Wound Healing: PerspectiveHanaNo ratings yet
- Anatomy, Physiology and Biochemistry of Cornea: Presenter-Yordanos (R1) Moderator-Dr YaredDocument106 pagesAnatomy, Physiology and Biochemistry of Cornea: Presenter-Yordanos (R1) Moderator-Dr YaredAmare GetnetNo ratings yet
- Library DissertationDocument201 pagesLibrary Dissertationhaneefmdf100% (2)
- The Epithelial Basement MembraneDocument9 pagesThe Epithelial Basement MembraneSitipradyta KasimNo ratings yet
- Cell Wall Layers: 1. Middle LamellaDocument11 pagesCell Wall Layers: 1. Middle Lamellafathimathshamla pkNo ratings yet
- Integumentary System: Stratum BasaleDocument21 pagesIntegumentary System: Stratum BasalerinaNo ratings yet
- Stratified Epithelia L 2Document45 pagesStratified Epithelia L 2srzvzvt7rzNo ratings yet
- 5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)Document115 pages5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)MobarobberNo ratings yet
- A4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Document16 pagesA4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Daniel CubasNo ratings yet
- SKINnerDocument9 pagesSKINnerjuleNo ratings yet
- Oral Mucous Membrane: DR Saurabh Juneja Professor and Head Dept of Oral Pathology SBBDCDocument54 pagesOral Mucous Membrane: DR Saurabh Juneja Professor and Head Dept of Oral Pathology SBBDCPuneet KaurNo ratings yet
- Natomy and Physiology of Cornea: Dr.K.Mahalakshmi 1 Yr PGDocument61 pagesNatomy and Physiology of Cornea: Dr.K.Mahalakshmi 1 Yr PGMaha lakshmiNo ratings yet
- Biology of Oral Mucosa and EsophagusDocument9 pagesBiology of Oral Mucosa and EsophagusJuan Sierra ZambranoNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- The 3 Layers of Skin: Epidermis, Dermis and Subcutaneous FatDocument11 pagesThe 3 Layers of Skin: Epidermis, Dermis and Subcutaneous FatPanggih Sekar Palupi IINo ratings yet
- Ex Vivo Expansion of Limbal Epithelial Stem Cells - Amniotic Membrane Serving As A Stem Cell NicheDocument16 pagesEx Vivo Expansion of Limbal Epithelial Stem Cells - Amniotic Membrane Serving As A Stem Cell NicheselcanekicierNo ratings yet
- Histology of Oral Mucous Membrane and GingivaDocument86 pagesHistology of Oral Mucous Membrane and GingivahazeemmegahedNo ratings yet
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Anatomy and Properties of BambooDocument198 pagesAnatomy and Properties of Bambooaya1604No ratings yet
- محاضره 2 انسجهDocument44 pagesمحاضره 2 انسجهYahya Daham Zafeer SakhrNo ratings yet
- EPITHELIUMDocument38 pagesEPITHELIUMAstridNo ratings yet
- ORAL MUCOSA NON-KERATINIZED EPITHELIUMDocument29 pagesORAL MUCOSA NON-KERATINIZED EPITHELIUMAMIT GUPTANo ratings yet
- Cell Spreading Kinetics Guided by Interfacial StiffnessDocument11 pagesCell Spreading Kinetics Guided by Interfacial StiffnesssaawanNo ratings yet
- Balda. Tight JDocument8 pagesBalda. Tight JrikaNo ratings yet
- Oral Mucous Membrane: by - Arindam MondalDocument92 pagesOral Mucous Membrane: by - Arindam MondalMohammed hisham khanNo ratings yet
- Oral Mucous Membrane: by - Arindam MondalDocument92 pagesOral Mucous Membrane: by - Arindam MondalMohammed hisham khanNo ratings yet
- The Three-dimensional Structure of the Cytostome-cytopharynxDocument11 pagesThe Three-dimensional Structure of the Cytostome-cytopharynxUNIG Aluana SantanaNo ratings yet
- Repaso Histologi ADocument132 pagesRepaso Histologi AMichael Rivera MoralesNo ratings yet
- Carcinoma Invasion and Metastasis A Role For EpithelialDocument5 pagesCarcinoma Invasion and Metastasis A Role For Epithelialfranciscrick69No ratings yet
- A Closer Look: The Corneal EpitheliumDocument4 pagesA Closer Look: The Corneal Epitheliumshay sNo ratings yet
- 11 B IntegumenttextDocument41 pages11 B IntegumenttextAbu Ibtihal OsmanNo ratings yet
- Epidermal and Dermal Adhesion StructuresDocument30 pagesEpidermal and Dermal Adhesion StructuresS FznsNo ratings yet
- Ijos 201346Document9 pagesIjos 201346Arnasary KaharNo ratings yet
- 2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisDocument16 pages2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisskbucherNo ratings yet
- Basic Physiology of The SkinDocument4 pagesBasic Physiology of The SkinRegina SinagaNo ratings yet
- 1.Integumentary System_histologyDocument34 pages1.Integumentary System_histologyqty9jgkpnzNo ratings yet
- Week 4 Case StudyDocument6 pagesWeek 4 Case StudyElizabeth GithinjiNo ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- 1 s2.0 S0022202X15447736 MainDocument7 pages1 s2.0 S0022202X15447736 MainKhan FatemaNo ratings yet
- Study of Cellwall Constituents and Cell InclusionsDocument42 pagesStudy of Cellwall Constituents and Cell Inclusionsharshe v70% (10)
- Hierarchical Levels of Organization of The BrazilDocument14 pagesHierarchical Levels of Organization of The BrazilwhaklNo ratings yet
- GIT Barrier Function JPDocument5 pagesGIT Barrier Function JPAhmed HamdyNo ratings yet
- GingivaDocument28 pagesGingivaAlvira MonicaNo ratings yet
- Extensive Squamous Metaplasia in Minor Salivary Gland PADocument7 pagesExtensive Squamous Metaplasia in Minor Salivary Gland PAGail Domecq TanawitNo ratings yet
- 2 Basal Lateral Apical modifications and Connective tissueDocument84 pages2 Basal Lateral Apical modifications and Connective tissueDeniz asmazNo ratings yet
- The Anatomy of The LimbusDocument8 pagesThe Anatomy of The LimbusBukaNo ratings yet
- نسخة نسخة نسخة ريميDocument6 pagesنسخة نسخة نسخة ريميAhmed QahtaniNo ratings yet
- Keratinisation and CornificationDocument38 pagesKeratinisation and Cornificationlizamjen100% (3)
- Anatomy and PhysiologyDocument25 pagesAnatomy and PhysiologyNaidin Catherine De Guzman-AlcalaNo ratings yet
- Cell Junctions: Specialized Cell Junctions Occur at Points of Cell-Cell andDocument8 pagesCell Junctions: Specialized Cell Junctions Occur at Points of Cell-Cell andRahul SharmaNo ratings yet
- Dentogingival Junction AnatomyDocument7 pagesDentogingival Junction AnatomyDrKrishna Das100% (1)
- Cellular Plasma Membrane Domains: Michael SheetzDocument3 pagesCellular Plasma Membrane Domains: Michael SheetzHonu Fran SilvaNo ratings yet
- Structure and Function of Membrane Proteins: Proceedings of the International Symposium on Structure and Function of Membrane Proteins Held in Selva Di Fasano (Italy), May 23-26, 1983From EverandStructure and Function of Membrane Proteins: Proceedings of the International Symposium on Structure and Function of Membrane Proteins Held in Selva Di Fasano (Italy), May 23-26, 1983E. QuagliarielloNo ratings yet
- MDP 39015006442878 57 1632815325Document1 pageMDP 39015006442878 57 1632815325Zachary DuongNo ratings yet
- MDP 39015006442878 49 1632815281Document1 pageMDP 39015006442878 49 1632815281Zachary DuongNo ratings yet
- MDP 39015006442878 41 1632815742Document1 pageMDP 39015006442878 41 1632815742Zachary DuongNo ratings yet
- Designing Undercut Brought Point The Pencil - Mark, LineDocument1 pageDesigning Undercut Brought Point The Pencil - Mark, LineZachary DuongNo ratings yet
- MDP 39015006442878 51 1632815293Document1 pageMDP 39015006442878 51 1632815293Zachary DuongNo ratings yet
- MDP 39015006442878 53 1632815303Document1 pageMDP 39015006442878 53 1632815303Zachary DuongNo ratings yet
- MDP 39015006442878 38 1632815715Document1 pageMDP 39015006442878 38 1632815715Zachary DuongNo ratings yet
- MDP 39015006442878 44 1632815770Document1 pageMDP 39015006442878 44 1632815770Zachary DuongNo ratings yet
- MDP 39015006442878 62 1632815350Document1 pageMDP 39015006442878 62 1632815350Zachary DuongNo ratings yet
- MDP 39015006442878 50 1632815287Document1 pageMDP 39015006442878 50 1632815287Zachary DuongNo ratings yet
- Two Flexi Tips Denture: or TrussDocument1 pageTwo Flexi Tips Denture: or TrussZachary DuongNo ratings yet
- Action For Bicus Lingual: and To End EndDocument1 pageAction For Bicus Lingual: and To End EndZachary DuongNo ratings yet
- MDP 39015006442878 65 1632815364Document1 pageMDP 39015006442878 65 1632815364Zachary DuongNo ratings yet
- MDP 39015006442878 59 1632815334Document1 pageMDP 39015006442878 59 1632815334Zachary DuongNo ratings yet
- MDP 39015006442878 46 1632815779Document1 pageMDP 39015006442878 46 1632815779Zachary DuongNo ratings yet
- MDP 39015006442878 47 1632815268Document1 pageMDP 39015006442878 47 1632815268Zachary DuongNo ratings yet
- MDP 39015006442878 43 1632815761Document1 pageMDP 39015006442878 43 1632815761Zachary DuongNo ratings yet
- MDP 39015006442878 23 1632815584Document1 pageMDP 39015006442878 23 1632815584Zachary DuongNo ratings yet
- MDP 39015006442878 21 1632815571Document1 pageMDP 39015006442878 21 1632815571Zachary DuongNo ratings yet
- MDP 39015006442878 45 1632816053Document1 pageMDP 39015006442878 45 1632816053Zachary DuongNo ratings yet
- MDP 39015006442878 22 1632815578Document1 pageMDP 39015006442878 22 1632815578Zachary DuongNo ratings yet
- MDP 39015006442878 39 1632815724Document1 pageMDP 39015006442878 39 1632815724Zachary DuongNo ratings yet
- MDP 39015006442878 37 1632815706Document1 pageMDP 39015006442878 37 1632815706Zachary DuongNo ratings yet
- MDP 39015006442878 60 1632815340Document1 pageMDP 39015006442878 60 1632815340Zachary DuongNo ratings yet
- Func Oc: Undercut Not Partial PartialDocument1 pageFunc Oc: Undercut Not Partial PartialZachary DuongNo ratings yet
- MDP 39015006442878 56 1632815320Document1 pageMDP 39015006442878 56 1632815320Zachary DuongNo ratings yet
- MDP 39015006442878 63 1632815355Document1 pageMDP 39015006442878 63 1632815355Zachary DuongNo ratings yet
- MDP 39015006442878 10 1632815446Document1 pageMDP 39015006442878 10 1632815446Zachary DuongNo ratings yet
- Will: Molar Lingual and Be Found Lingual of Buccal ofDocument1 pageWill: Molar Lingual and Be Found Lingual of Buccal ofZachary DuongNo ratings yet
- MDP 39015006442878 29 1632815625Document1 pageMDP 39015006442878 29 1632815625Zachary DuongNo ratings yet
- The Four Major Organ Systems of the Human BodyDocument5 pagesThe Four Major Organ Systems of the Human BodyXyramhel AcirolNo ratings yet
- 7 OvuleDocument6 pages7 OvuleAlexandra Maria NeaguNo ratings yet
- Ornitz, 1974 - The Modulation Os Sensory Input and Motor Output in Autistic ChildrenDocument19 pagesOrnitz, 1974 - The Modulation Os Sensory Input and Motor Output in Autistic ChildrenRaisa CoppolaNo ratings yet
- SREO BiologyDocument8 pagesSREO BiologySaktheeswaran VNo ratings yet
- Clinical Hematology and Fundamentals of Hemostasis 5th Edition Harmening Test BankDocument36 pagesClinical Hematology and Fundamentals of Hemostasis 5th Edition Harmening Test Bankgeincupola.06zi100% (23)
- Endometriosis An Ayurvedic ViewDocument7 pagesEndometriosis An Ayurvedic ViewroykibetNo ratings yet
- Lesson Plan:: Teacher Name: Grade: SubjectDocument3 pagesLesson Plan:: Teacher Name: Grade: SubjectHaifa Mohammadsali AndilingNo ratings yet
- Anatomy and Physiology of The Digestive SystemDocument5 pagesAnatomy and Physiology of The Digestive SystemKate Cris TalatagodNo ratings yet
- Physio Ex Act 6Document11 pagesPhysio Ex Act 6alifia azzahraNo ratings yet
- Examination BiologyDocument3 pagesExamination BiologyKosh MatthewNo ratings yet
- Gastrointestinal PhysiologyDocument39 pagesGastrointestinal PhysiologyKaty SanchezNo ratings yet
- CP-Guillain Barre SyndromeDocument35 pagesCP-Guillain Barre Syndromekathy60% (5)
- Darah Dan Fungsi DarahDocument14 pagesDarah Dan Fungsi DarahdelisNo ratings yet
- Hydra 1Document25 pagesHydra 1Gowtham GupthaNo ratings yet
- Who-Is-It Teacher SwitcherDocument21 pagesWho-Is-It Teacher SwitcherАлина СолонскаяNo ratings yet
- Soft Tissue Tumor Treatment and Diagnosis in the NeckDocument15 pagesSoft Tissue Tumor Treatment and Diagnosis in the NeckCynthia BarrientosNo ratings yet
- Greek Latin Roots AssignmentDocument4 pagesGreek Latin Roots Assignmentapi-241263980No ratings yet
- Male Reproductive Anatomy and FunctionDocument42 pagesMale Reproductive Anatomy and FunctionBlodin ZylfiuNo ratings yet
- Fine Needle Aspiration Cytology (FNAC) of GISTDocument7 pagesFine Needle Aspiration Cytology (FNAC) of GISTurfriendanshul100% (1)
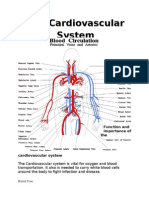
- Function and Importance of The Cardiovascular SystemDocument3 pagesFunction and Importance of The Cardiovascular SystemBenjamin SmithNo ratings yet
- Tetanus PathoDocument3 pagesTetanus PathoElisha Gine AndalesNo ratings yet
- Nanobiotechnology-Based Biosensors For Rapid Detection of Bacterial ContaminantsDocument59 pagesNanobiotechnology-Based Biosensors For Rapid Detection of Bacterial ContaminantsRachel PlanillaNo ratings yet
- Circulation & HemodynamicsDocument7 pagesCirculation & HemodynamicsFaria Islam JuhiNo ratings yet
- Congenital SyphilisDocument3 pagesCongenital SyphilisLakshya J BasumataryNo ratings yet
- Haematology Diagnosis-Army USADocument205 pagesHaematology Diagnosis-Army USAManuela BotisNo ratings yet
- Parasympathetic Nervous SystemDocument20 pagesParasympathetic Nervous SystemShahid Maqbool100% (1)
- Far Eastern University Institue of Arts and Science Department of Medical Technology Histology 2 SEMESTER AY 2020-2021Document1 pageFar Eastern University Institue of Arts and Science Department of Medical Technology Histology 2 SEMESTER AY 2020-2021Kaye RiofloridoNo ratings yet
- Deep Venous Thrombosis: Practice EssentialsDocument27 pagesDeep Venous Thrombosis: Practice Essentialsphoechoex100% (1)
- Year 8 Science OverviewDocument4 pagesYear 8 Science Overviewapi-3137245670% (1)
- Anatomy and PhysiologyDocument12 pagesAnatomy and PhysiologyGregg PalabricaNo ratings yet