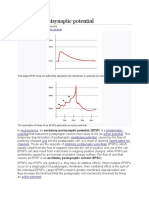
You might also like
- Chulbae 1996Document15 pagesChulbae 1996M4shroomNo ratings yet
- BCR 1976Document15 pagesBCR 1976Tia YuandaNo ratings yet
- Propriospinal Control of Interneurons inDocument4 pagesPropriospinal Control of Interneurons inAgus WiyonoNo ratings yet
- TMP F204Document2 pagesTMP F204FrontiersNo ratings yet
- (Journal of Neurosurgery) Electrophysiological Connections Between The Hippocampus and Entorhinal Cortex in Patients With Complex Partial SeizuresDocument9 pages(Journal of Neurosurgery) Electrophysiological Connections Between The Hippocampus and Entorhinal Cortex in Patients With Complex Partial SeizuresJosé luis Capellini donathNo ratings yet
- Laryngeal Synergy in Singing: Jéécam 'Eic#Evtd, EewDocument6 pagesLaryngeal Synergy in Singing: Jéécam 'Eic#Evtd, EewPaulaRiveroNo ratings yet
- Orexin Neuron Activity in Mating Mice - A Pilot STDocument6 pagesOrexin Neuron Activity in Mating Mice - A Pilot STRika LauraNo ratings yet
- Brain, Breathing and Breathless PDFDocument8 pagesBrain, Breathing and Breathless PDFJesus Laserna RuizNo ratings yet
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickNo ratings yet
- Inhibición y Curvas de Sintonía en NCV - FullDocument14 pagesInhibición y Curvas de Sintonía en NCV - FullSilvio SánchezNo ratings yet
- Types of Neurons in The Enteric Nervous SystemDocument10 pagesTypes of Neurons in The Enteric Nervous SystemErnesto Ochoa MonroyNo ratings yet
- Ader, Cohen, 1982 - Behaviorally Conditioned Immunosuppresion and Murine Systemic Lupus ErythematosuDocument3 pagesAder, Cohen, 1982 - Behaviorally Conditioned Immunosuppresion and Murine Systemic Lupus ErythematosuDiogo Deganeli de Brito VaragoNo ratings yet
- Braistem ReflectsDocument9 pagesBraistem ReflectsPedro Waskar Aramayo TorricoNo ratings yet
- Aids To The Examination of The Peripheral Nervous System - MRC Memorandum No.45 (Superseding War Memorandum No.7) PDFDocument70 pagesAids To The Examination of The Peripheral Nervous System - MRC Memorandum No.45 (Superseding War Memorandum No.7) PDFDanielNo ratings yet
- NIH Public Access: Author ManuscriptDocument13 pagesNIH Public Access: Author ManuscriptRoar SyltebøNo ratings yet
- Seminario 2Document20 pagesSeminario 2LeonardoNo ratings yet
- Placement of Probes in Electrostimulation and Biofeedback Training in Pelvic Floor DysfunctionDocument6 pagesPlacement of Probes in Electrostimulation and Biofeedback Training in Pelvic Floor DysfunctionCorey EasleyNo ratings yet
- Gundlfinger Et Al. - 2010 - Natural Spike Trains Trigger Short - and Long Lasting Dynamics at Hippocampal Mossy Fiber Synapses in RodentsDocument9 pagesGundlfinger Et Al. - 2010 - Natural Spike Trains Trigger Short - and Long Lasting Dynamics at Hippocampal Mossy Fiber Synapses in RodentsGabriel HerreraNo ratings yet
- Electroencephalography (Eeg) : Atarzyna Linowska Iotr Urka Warsaw University Warszawa, PolandDocument15 pagesElectroencephalography (Eeg) : Atarzyna Linowska Iotr Urka Warsaw University Warszawa, PolandNicholas DiazNo ratings yet
- 2000 Brain ALSDocument16 pages2000 Brain ALSSultan TarlacıNo ratings yet
- Takuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1Document18 pagesTakuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1FedrmNo ratings yet
- 10 1016@j Aanat 2019 151439Document40 pages10 1016@j Aanat 2019 151439CFS HyderabadNo ratings yet
- Bear&KirkwoodDocument11 pagesBear&KirkwoodswagatarcNo ratings yet
- Gibbins2009 PDFDocument1 pageGibbins2009 PDFDR GANo ratings yet
- Abdominal Motor UnityDocument9 pagesAbdominal Motor UnityBrenda Soares Nogueira BritoNo ratings yet
- Vertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsDocument7 pagesVertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsFractalScribd707No ratings yet
- Cap 11 - 22 Handbook of Clinical AudiologyDocument216 pagesCap 11 - 22 Handbook of Clinical AudiologyCami IrribarraNo ratings yet
- Acta HumanDocument8 pagesActa HumanjoaomonterosNo ratings yet
- Plasticity in The Human Central Nervous SystemDocument15 pagesPlasticity in The Human Central Nervous SystemMarcelo LugonNo ratings yet
- Anaesthesia - October 1987 - Norley - Erroneous actuation of the pulse oximeter (1)Document1 pageAnaesthesia - October 1987 - Norley - Erroneous actuation of the pulse oximeter (1)hayaatmueen45No ratings yet
- Morales1978 AtoniaDocument7 pagesMorales1978 AtoniasujNo ratings yet
- Topological Distribution of Oddball P300' ResponsesDocument8 pagesTopological Distribution of Oddball P300' ResponsesAhmad Fathy MahmoudNo ratings yet
- Harry Shpuntoff, and William Shpuntoff,: Jackson Heights, N. YDocument8 pagesHarry Shpuntoff, and William Shpuntoff,: Jackson Heights, N. YSatya AsatyaNo ratings yet
- Hirudo Medicinalis Local Bending: Myomodulin Effects On The P To AP SynapseDocument10 pagesHirudo Medicinalis Local Bending: Myomodulin Effects On The P To AP SynapseIlavarasi PanduranganNo ratings yet
- Husos de SueñoDocument37 pagesHusos de SueñoYair FloresNo ratings yet
- Of Of: The Role The System in Phenomena Temporal Lobe EpilepsyDocument16 pagesOf Of: The Role The System in Phenomena Temporal Lobe EpilepsyBONA VICTOR SIMANJUNTAK MDNo ratings yet
- Anterior Nerve Syndrome - Morton SpinnerDocument11 pagesAnterior Nerve Syndrome - Morton SpinnerJunji Miller FukuyamaNo ratings yet
- Short Communication Sleep Disturbances in A Case of Brain-Stem Lesions Pharmacological StudyDocument3 pagesShort Communication Sleep Disturbances in A Case of Brain-Stem Lesions Pharmacological StudymorexyzNo ratings yet
- Aculputura P InferDocument5 pagesAculputura P InferRhayane CoelhoNo ratings yet
- Delorme 1997Document8 pagesDelorme 1997groot marvelNo ratings yet
- K-Complexes and Sleep Spindles Before Transient Activation During SleepDocument8 pagesK-Complexes and Sleep Spindles Before Transient Activation During SleepHernán MarínNo ratings yet
- Uniiateral Forced Nostril Breathing:: Basic Science, Clinical Trials, and Selected Advanced TechniquesDocument28 pagesUniiateral Forced Nostril Breathing:: Basic Science, Clinical Trials, and Selected Advanced TechniquesCambiador de MundoNo ratings yet
- Rozzi EJN2008subrayadoDocument20 pagesRozzi EJN2008subrayadoMaria Elena Ceballos VillegasNo ratings yet
- Woelfel. Electromyographic Evidence Suporting The Mandibular Hinge Axis Theoy. (1957)Document7 pagesWoelfel. Electromyographic Evidence Suporting The Mandibular Hinge Axis Theoy. (1957)Jose Enrique AvilaNo ratings yet
- T.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigDocument18 pagesT.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigLuammmNo ratings yet
- Experimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalDocument13 pagesExperimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalAndré DiasNo ratings yet
- Lab 02 Exciteable tissue_Student F2020Document14 pagesLab 02 Exciteable tissue_Student F2020Alena JosephNo ratings yet
- tmp5703 TMPDocument11 pagestmp5703 TMPFrontiersNo ratings yet
- 1981 L-To 3rd Order J Comp PhysiolDocument12 pages1981 L-To 3rd Order J Comp PhysiolClub OrilyonNo ratings yet
- Tasa de Transporte AxoplasmicoDocument19 pagesTasa de Transporte AxoplasmicoAndy Moreno GalvezNo ratings yet
- Jphysiol00347 0056Document13 pagesJphysiol00347 0056MichaelNo ratings yet
- SSEPDocument12 pagesSSEPahippo100% (1)
- Excitatory Postsynaptic Potential: Jump To Navigation Jump To SearchDocument6 pagesExcitatory Postsynaptic Potential: Jump To Navigation Jump To SearchJennie KimNo ratings yet
- EMG Lead PlacementDocument14 pagesEMG Lead PlacementalanNo ratings yet
- Resume NesDocument4 pagesResume NesAlexandra FodorNo ratings yet
- RAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeDocument14 pagesRAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeFedrmNo ratings yet
- Sympathetic Skin Response in Obstructive Sleep Apnea SyndromeDocument9 pagesSympathetic Skin Response in Obstructive Sleep Apnea SyndromeGyörgy BáthoriNo ratings yet
- Jphysiol00385 0110Document9 pagesJphysiol00385 0110MichaelNo ratings yet
- Carr Central ProjectionsDocument13 pagesCarr Central ProjectionsMengda ZhangNo ratings yet
- Plasticity of Connections Underlying LocDocument25 pagesPlasticity of Connections Underlying LocAgus WiyonoNo ratings yet
- ENEURO 0004-14 2014 FullDocument33 pagesENEURO 0004-14 2014 FullnewhaberNo ratings yet
- Cleland and Madhavan 2021 - Ipsilateral Motor Pathways To The Lower Limb After Stroke - Insights and OpportunitiesDocument14 pagesCleland and Madhavan 2021 - Ipsilateral Motor Pathways To The Lower Limb After Stroke - Insights and OpportunitiesAgus WiyonoNo ratings yet
- Step Training Dependent Plasticity in SPDocument11 pagesStep Training Dependent Plasticity in SPAgus WiyonoNo ratings yet
- Cleland and Madhavan 2021 - Ipsilateral Motor Pathways To The Lower Limb After Stroke - Insights and OpportunitiesDocument14 pagesCleland and Madhavan 2021 - Ipsilateral Motor Pathways To The Lower Limb After Stroke - Insights and OpportunitiesAgus WiyonoNo ratings yet
- Immediate effects of therapeutic facilitation on hemiparetic gaitDocument8 pagesImmediate effects of therapeutic facilitation on hemiparetic gaitAgus WiyonoNo ratings yet
- Best Core Stabilization To Subcortical PlasticityDocument7 pagesBest Core Stabilization To Subcortical PlasticityAgus WiyonoNo ratings yet
- Bio Marker of Stroke Recovery Boyd2017Document14 pagesBio Marker of Stroke Recovery Boyd2017Agus WiyonoNo ratings yet
- Bobath Concept Structural Framework (BCSF) : Positioning Partial Aspects Within A Holistic Therapeutic ConceptDocument7 pagesBobath Concept Structural Framework (BCSF) : Positioning Partial Aspects Within A Holistic Therapeutic ConceptekaNo ratings yet
- Best Core Stabilization To Subcortical PlasticityDocument7 pagesBest Core Stabilization To Subcortical PlasticityAgus WiyonoNo ratings yet
- Adaptive Plasticity and Recovery 2014DaliseArchItalBiolDocument27 pagesAdaptive Plasticity and Recovery 2014DaliseArchItalBiolAgus WiyonoNo ratings yet
- Bobath Concept Structural Framework (BCSF) : Positioning Partial Aspects Within A Holistic Therapeutic ConceptDocument7 pagesBobath Concept Structural Framework (BCSF) : Positioning Partial Aspects Within A Holistic Therapeutic ConceptekaNo ratings yet
- Bio Marker of Stroke Recovery Boyd2017Document14 pagesBio Marker of Stroke Recovery Boyd2017Agus WiyonoNo ratings yet
- Lowbackpaindecembersuhash F 110118024849 Phpapp01Document39 pagesLowbackpaindecembersuhash F 110118024849 Phpapp01Agus WiyonoNo ratings yet
- Rooij Et Al-2016-Arthritis Care & ResearchDocument12 pagesRooij Et Al-2016-Arthritis Care & ResearchJennifer JaneNo ratings yet
- Adaptive Plasticity and Recovery 2014DaliseArchItalBiolDocument27 pagesAdaptive Plasticity and Recovery 2014DaliseArchItalBiolAgus WiyonoNo ratings yet
- Clinical Applications of DTI and Fiber Tractography LeeDocument40 pagesClinical Applications of DTI and Fiber Tractography LeeAgus WiyonoNo ratings yet
- Matecconf - Icet4sd2018 - 01083 Tugas Penunjang Diagnostik 1Document6 pagesMatecconf - Icet4sd2018 - 01083 Tugas Penunjang Diagnostik 1Agus WiyonoNo ratings yet
- Brain Changes Following 4 Weeks Trainingsale2017Document15 pagesBrain Changes Following 4 Weeks Trainingsale2017Agus WiyonoNo ratings yet
- Functional Reach Test ProtocolDocument3 pagesFunctional Reach Test Protocolwenda rosita setyaningsihNo ratings yet
- Jurnal RadiologiDocument9 pagesJurnal RadiologiAgus WiyonoNo ratings yet
- Management of Osteoporosis and Associated QOL in Post Menopausal WomenDocument12 pagesManagement of Osteoporosis and Associated QOL in Post Menopausal WomenHolubiac Iulian StefanNo ratings yet
- NIH Public Access: Exercise-Enhanced Neuroplasticity Targeting Motor and Cognitive Circuitry in Parkinson's DiseaseDocument19 pagesNIH Public Access: Exercise-Enhanced Neuroplasticity Targeting Motor and Cognitive Circuitry in Parkinson's DiseaseAgus WiyonoNo ratings yet
- Action PotentialDocument16 pagesAction PotentialAgus WiyonoNo ratings yet
- OaDocument6 pagesOaAgus WiyonoNo ratings yet
- QiGong Global SummitDocument45 pagesQiGong Global SummittoanomisNo ratings yet
- Speech Mechanism 1aDocument30 pagesSpeech Mechanism 1aNathania AnindyaNo ratings yet
- Optimization of Granulation and Compression Process Variables of Atenolol Tablets Using Box Behnken DesignDocument9 pagesOptimization of Granulation and Compression Process Variables of Atenolol Tablets Using Box Behnken DesignnkszoneNo ratings yet
- SulfapiridinaDocument21 pagesSulfapiridinaAylinNayeliNo ratings yet
- Daftar Pustaka Case CephalgiaDocument3 pagesDaftar Pustaka Case Cephalgiakatherine rinovaNo ratings yet
- Pediatric Fever, Cough and Rash ExamDocument3 pagesPediatric Fever, Cough and Rash ExamTheju ReddyNo ratings yet
- SYLABUSDocument6 pagesSYLABUSLarimar Gonzalez LoraNo ratings yet
- HospDocument414 pagesHospjnaguNo ratings yet
- Pt. Indra Giri Raya: Kode Nama Barang Stock Harga Jual SatuanDocument12 pagesPt. Indra Giri Raya: Kode Nama Barang Stock Harga Jual Satuanrema_rhere95No ratings yet
- ParkMediclaim Insurance TPA PVT LTD Group Policy Search - KarnatakaDocument21 pagesParkMediclaim Insurance TPA PVT LTD Group Policy Search - KarnatakakumarNo ratings yet
- Sample Exhibit FormDocument14 pagesSample Exhibit FormCharm BoiserNo ratings yet
- Kate Laver's Primary FRCA Guide: My Revision (For Exam in June)Document3 pagesKate Laver's Primary FRCA Guide: My Revision (For Exam in June)M H DuNo ratings yet
- ReadingDocument19 pagesReadingXuyến Chi TrầnNo ratings yet
- Alexander DisciplineDocument240 pagesAlexander DisciplineOctavian Tavi100% (1)
- Orgone Diffuser Reiki GuideDocument7 pagesOrgone Diffuser Reiki GuideAndrea Koumarian100% (1)
- A Devoted SonDocument16 pagesA Devoted SonKirran Khumar Golla50% (4)
- 3rd Year MUHS Winter 2022Document10 pages3rd Year MUHS Winter 2022Juned LabbaiNo ratings yet
- Drugs Affecting Neurologic SystemDocument5 pagesDrugs Affecting Neurologic SystemjustineGREATNo ratings yet
- Challenges in Implantable BioMEMSDocument8 pagesChallenges in Implantable BioMEMSConstantino Miguelez PeñaNo ratings yet
- Manus 2022Document60 pagesManus 2022Benevolentia Foundation100% (4)
- Mentorship FinalDocument46 pagesMentorship Finalapi-200572195No ratings yet
- Antigen Antibody ReactionDocument16 pagesAntigen Antibody ReactionNadzierah RazakNo ratings yet
- Reiki Tips You Haven T Thought ofDocument35 pagesReiki Tips You Haven T Thought ofAnonymous 3gHvQv100% (3)
- Use of Animals in Product DevelopmentDocument4 pagesUse of Animals in Product DevelopmentMerih SunNo ratings yet
- Functional Analysis and Treatment of Inappropriate Verbal BehaviorDocument3 pagesFunctional Analysis and Treatment of Inappropriate Verbal BehaviorMariaNo ratings yet
- White Sponge NevusDocument2 pagesWhite Sponge Nevusbundahara zoomNo ratings yet
- High Alert Medication PlanDocument15 pagesHigh Alert Medication PlanNina Morada100% (5)
- Delta Sigma Theta Sorority, Inc. Jacksonville Alumnae ChapterDocument7 pagesDelta Sigma Theta Sorority, Inc. Jacksonville Alumnae ChaptermortensenkNo ratings yet
- Synthetic Antibacterial AgentsDocument58 pagesSynthetic Antibacterial AgentsApurba Sarker Apu100% (3)