You might also like
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsFrom EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNo ratings yet
- Yuandani 2013Document10 pagesYuandani 2013Khrisna Whaty SilalahiNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- Zii2043 PDFDocument10 pagesZii2043 PDFPablo VelazquezNo ratings yet
- Cimid 2015 AjayDocument7 pagesCimid 2015 AjayprashantcdriNo ratings yet
- Lin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesDocument15 pagesLin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesJanina PuenteNo ratings yet
- Patogenos e InmunidadDocument11 pagesPatogenos e InmunidadMagaly CarrilloNo ratings yet
- Molecular analysis of porin gene transcription in multidrug-resistant E. coliDocument10 pagesMolecular analysis of porin gene transcription in multidrug-resistant E. coliThreshNo ratings yet
- 1 s2.0 S0021925820870148 MainDocument11 pages1 s2.0 S0021925820870148 MainAina SarahNo ratings yet
- Cytokine Profiling of Macrophages Exposed To Porphyromonas Gingivalis, Its Lipopolysaccharide, or Its Fima ProteinDocument10 pagesCytokine Profiling of Macrophages Exposed To Porphyromonas Gingivalis, Its Lipopolysaccharide, or Its Fima Proteinapi-26071302No ratings yet
- Fungos Entomopatogenico Interação Hospedeiro e VetorDocument58 pagesFungos Entomopatogenico Interação Hospedeiro e VetorGabriel Henrique Santos SilveiraNo ratings yet
- Kamada2013 PDFDocument15 pagesKamada2013 PDFÂngela Souza BarroqueiroNo ratings yet
- Bacterial Expression and Enzymatic Activity Analysis of ME1 A Ribosome-Inactivating Protein From Mirabilis Expansa - USADocument10 pagesBacterial Expression and Enzymatic Activity Analysis of ME1 A Ribosome-Inactivating Protein From Mirabilis Expansa - USAT AmaruNo ratings yet
- Molecular Analysis of Antimicrobial Resistance Mechanisms in N-GonoDocument11 pagesMolecular Analysis of Antimicrobial Resistance Mechanisms in N-GonoMulatuNo ratings yet
- Phagocytosis in A Chemr23-And Chemerin Peptides Promote: Data SupplementaryDocument11 pagesPhagocytosis in A Chemr23-And Chemerin Peptides Promote: Data Supplementarybiosynthesis12No ratings yet
- The Immunomodulatory Activities of Picria Fel-TerrDocument5 pagesThe Immunomodulatory Activities of Picria Fel-TerrMuhammad Fauzan LubisNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
- The Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Document7 pagesThe Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Cristian Gutiérrez VeraNo ratings yet
- Fimmu 06 00555Document20 pagesFimmu 06 00555www.henry1020No ratings yet
- Enteric bacteria boost defences against oxidative stress in Entamoeba histolyticaDocument12 pagesEnteric bacteria boost defences against oxidative stress in Entamoeba histolyticagwyneth.green.512No ratings yet
- Microbial Virulence Factors: Molecular SciencesDocument6 pagesMicrobial Virulence Factors: Molecular SciencesAnasNo ratings yet
- Differential Host Pro-Inflammatory ResponseDocument10 pagesDifferential Host Pro-Inflammatory ResponseMohammad Asef HgNo ratings yet
- Evaluation of Classical, Alternative, and Regulatory Functions of Bone Marrow-Derived MacrophagesDocument11 pagesEvaluation of Classical, Alternative, and Regulatory Functions of Bone Marrow-Derived MacrophagesgugicevdzoceNo ratings yet
- β-Trefoil Protease Inhibitors from Higher FungiDocument9 pagesβ-Trefoil Protease Inhibitors from Higher FungiGregorius HocevarNo ratings yet
- Mitochondrial Complex 1is Important For Plant Tolerance To Fungal Biotic StressDocument11 pagesMitochondrial Complex 1is Important For Plant Tolerance To Fungal Biotic StressSryahwa PublicationsNo ratings yet
- Inadequate Timing Between Corticosteroid and Antibiotics Applications Increases Mortality Due To SepsisDocument7 pagesInadequate Timing Between Corticosteroid and Antibiotics Applications Increases Mortality Due To SepsisDwi afrianti Rahma putriNo ratings yet
- Journal Pone 0274005Document20 pagesJournal Pone 0274005Kim VănNo ratings yet
- Fimmu 11 571816Document20 pagesFimmu 11 571816Ricardo GomezNo ratings yet
- 1 s2.0 S1931312815004990 MainDocument13 pages1 s2.0 S1931312815004990 MainNouman SandhuNo ratings yet
- Popov 2002Document7 pagesPopov 2002DekdesNo ratings yet
- Dayakar2015. Evaluacion Del Efecto Inmunomodulador in Vitro e in VivoDocument10 pagesDayakar2015. Evaluacion Del Efecto Inmunomodulador in Vitro e in VivoLaura GarciaNo ratings yet
- Viale 2014Document9 pagesViale 2014Maria Laura MonNo ratings yet
- G 2.1 Chemotaxis-BacteriaDocument8 pagesG 2.1 Chemotaxis-BacteriaDiego LujanNo ratings yet
- In Silico Characterization of Meloidogyne Genus: Nematode Cellulose Binding ProteinsDocument13 pagesIn Silico Characterization of Meloidogyne Genus: Nematode Cellulose Binding ProteinsyutefupNo ratings yet
- Singh 2010Document8 pagesSingh 2010bibek1306No ratings yet
- Jir2018 6529681Document6 pagesJir2018 6529681Herry JapariNo ratings yet
- Parasites Evade TheDocument7 pagesParasites Evade TheMike zombieNo ratings yet
- Pathogen Strategies To Evade Innate Immune Response A Signaling Point of ViewDocument43 pagesPathogen Strategies To Evade Innate Immune Response A Signaling Point of Viewكرار جابرNo ratings yet
- Peptidoglycan Recognition Proteins Modulators of The Microbiome and InflammationDocument15 pagesPeptidoglycan Recognition Proteins Modulators of The Microbiome and Inflammationdavid onyangoNo ratings yet
- J. Biol. Chem.-2019-Coelho-1202-17 PDFDocument17 pagesJ. Biol. Chem.-2019-Coelho-1202-17 PDFJoel Torres VillenaNo ratings yet
- Functional Differences and Similarities in Activated Peripheral Blood Mononuclear Cells by Lipopolysaccharide or Phytohemagglutinin Stimulation Between Human and Cynomolgus Monkeys PDFDocument25 pagesFunctional Differences and Similarities in Activated Peripheral Blood Mononuclear Cells by Lipopolysaccharide or Phytohemagglutinin Stimulation Between Human and Cynomolgus Monkeys PDFLluis GomezNo ratings yet
- Prolactin Increase Frecuency of T Helper FolicularDocument15 pagesProlactin Increase Frecuency of T Helper Folicularauroragamez2015No ratings yet
- 2022 International Immunopharmacology Q1RamírezDocument15 pages2022 International Immunopharmacology Q1RamírezAndres RamirezNo ratings yet
- International ImmunopharmacologyDocument15 pagesInternational ImmunopharmacologyAndres RamirezNo ratings yet
- HDACDocument10 pagesHDACfarihameetsworldNo ratings yet
- Molecular Modelling, Docking and Interaction Studies of Human-Plasmogen and Salmonella-Enolase With Enolase InhibitorsDocument4 pagesMolecular Modelling, Docking and Interaction Studies of Human-Plasmogen and Salmonella-Enolase With Enolase Inhibitorskarthikeyan90No ratings yet
- 2020 Article 16120Document15 pages2020 Article 16120Matheus TavaresNo ratings yet
- SdarticleDocument7 pagesSdarticlenandhus2227No ratings yet
- Pi Is 0021925818816519Document7 pagesPi Is 0021925818816519Claudio IngrossoNo ratings yet
- Role of Elicitors in Activating Plant Defenses Against StressDocument9 pagesRole of Elicitors in Activating Plant Defenses Against StressVaibhav UpadhyayNo ratings yet
- Jun Zou, Nathan ShankarDocument11 pagesJun Zou, Nathan ShankarHQ HQNo ratings yet
- TMP 662Document11 pagesTMP 662FrontiersNo ratings yet
- Ec 00174-07Document8 pagesEc 00174-07dj1223549No ratings yet
- recognition -Document10 pagesrecognition -Sapna VermaNo ratings yet
- Virulence ToxinsDocument10 pagesVirulence ToxinsAvesh TiwariNo ratings yet
- Lig A LeptospiraDocument10 pagesLig A Leptospiraeduardorestrepo2000No ratings yet
- Molecules 28 02202Document16 pagesMolecules 28 02202Sandra Aliaga MorNo ratings yet
- Plasmodium Falciparum Apical Membrane Antigen 1 (Pfama1)Document1 pagePlasmodium Falciparum Apical Membrane Antigen 1 (Pfama1)Pamella CristinyNo ratings yet
- Revista Brasileira de ZootecniaDocument7 pagesRevista Brasileira de ZootecniayutefupNo ratings yet
- Immunomodulation in The Canine Endometrium by UteropathogenicDocument17 pagesImmunomodulation in The Canine Endometrium by UteropathogenicAndreea Blanco VeraNo ratings yet
- 2017-Justicia Adhatoda MidgutDocument11 pages2017-Justicia Adhatoda MidgutDwiNo ratings yet
- 2016-Inward Rectifier Potassium (Kir)Document10 pages2016-Inward Rectifier Potassium (Kir)DwiNo ratings yet
- 2020-Srikaya Seed Granule Different TemperatureDocument7 pages2020-Srikaya Seed Granule Different TemperatureDwiNo ratings yet
- PROSIDINGPOKJANASTOIDocument198 pagesPROSIDINGPOKJANASTOIPaundra DeniNo ratings yet
- 2016-Andrographolide Dari Andrographis PaniculataDocument39 pages2016-Andrographolide Dari Andrographis PaniculataDwiNo ratings yet
- 2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDocument7 pages2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDwiNo ratings yet
- A Toxin-Binding Alkaline PhospDocument20 pagesA Toxin-Binding Alkaline PhospDwiNo ratings yet
- A Transcriptomic Survey of TheDocument20 pagesA Transcriptomic Survey of TheDwiNo ratings yet
- 2014-Maybe Used-Ocotea CymbarumDocument10 pages2014-Maybe Used-Ocotea CymbarumDwiNo ratings yet
- Action of < I> MetarhiziumDocument10 pagesAction of < I> MetarhiziumDwiNo ratings yet
- A Transformed Bacterium ExpresDocument16 pagesA Transformed Bacterium ExpresDwiNo ratings yet
- Action of Natural PhytosanitarDocument10 pagesAction of Natural PhytosanitarDwiNo ratings yet
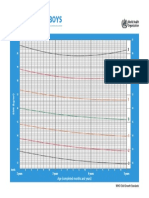
- CHT Bfa Boys Z 2 5Document1 pageCHT Bfa Boys Z 2 5DwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
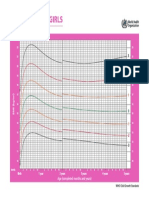
- CHT Bfa Girls Z 0 5Document1 pageCHT Bfa Girls Z 0 5DwiNo ratings yet
- Attachment 1706 631288cdc2e9fDocument4 pagesAttachment 1706 631288cdc2e9fDwiNo ratings yet
- Role of cytochrome P450s in insecticide resistanceDocument12 pagesRole of cytochrome P450s in insecticide resistanceDwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
- Attachment 1706 631288cdc288dDocument1 pageAttachment 1706 631288cdc288dDwiNo ratings yet
- Attachment 1706 631739d8303e3Document1 pageAttachment 1706 631739d8303e3DwiNo ratings yet
- ILMU PENYAKIT SARAF RSAU ESNAWAN ANTARIKSADocument5 pagesILMU PENYAKIT SARAF RSAU ESNAWAN ANTARIKSADwiNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- 2012-Denovo NilaparvataDocument9 pages2012-Denovo NilaparvataDwiNo ratings yet
- Plant Polypeptides Review: Fungitoxic and Insecticidal EffectsDocument18 pagesPlant Polypeptides Review: Fungitoxic and Insecticidal EffectsDwiNo ratings yet
- Euphorbia Extracts Impact on Moth DevelopmentDocument10 pagesEuphorbia Extracts Impact on Moth DevelopmentDwiNo ratings yet
- 2012-Drosophilla Melanogaster Metabolism InsecticideDocument7 pages2012-Drosophilla Melanogaster Metabolism InsecticideDwiNo ratings yet
- 2012-Garlic-Allium Sativum EndotoxinDocument11 pages2012-Garlic-Allium Sativum EndotoxinDwiNo ratings yet
- 2012-CYP450 CYP6B47 cDNAsDocument11 pages2012-CYP450 CYP6B47 cDNAsDwiNo ratings yet
- 2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDocument682 pages2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDwiNo ratings yet
- Onepot Xldle-2: SBQ Direct Emulsion Highly Sensitive Emulsion For DLE/LEDDocument1 pageOnepot Xldle-2: SBQ Direct Emulsion Highly Sensitive Emulsion For DLE/LEDerfan ahmedNo ratings yet
- Assignment # 2: Thermo LabDocument4 pagesAssignment # 2: Thermo LabmjunaidNo ratings yet
- Soil Test Interpretation EUROCODE 1999Document8 pagesSoil Test Interpretation EUROCODE 1999RanoulfoNo ratings yet
- Material SpecificationDocument10 pagesMaterial SpecificationMuthu GaneshNo ratings yet
- Echem WKST KeyDocument7 pagesEchem WKST KeyNurul Hana OmarNo ratings yet
- BiofertilizerDocument5 pagesBiofertilizerUmeshNo ratings yet
- REACTION RATE AND ACTIVATION ENERGY OF ETHYL ACETATEDocument6 pagesREACTION RATE AND ACTIVATION ENERGY OF ETHYL ACETATEGlënn Märk PrësörësNo ratings yet
- Flange IntegrityDocument46 pagesFlange IntegrityPaul PhiliphsNo ratings yet
- Guide No01 GRLs7th Issue Oct21Document13 pagesGuide No01 GRLs7th Issue Oct21hüseyinNo ratings yet
- Differences Between A Thermostatic Expansion Valve and An Electronic Expansion ValveDocument4 pagesDifferences Between A Thermostatic Expansion Valve and An Electronic Expansion Valvexuyen tran100% (1)
- Heteropolysaccharides Lecture For 1st Yr MBBS by DR Waseem KausarDocument20 pagesHeteropolysaccharides Lecture For 1st Yr MBBS by DR Waseem KausarIMDCBiochem67% (6)
- Quantify Vinegar AcidityDocument7 pagesQuantify Vinegar AcidityJoseline SorianoNo ratings yet
- Chapter 4-Aromatic CompoundsDocument48 pagesChapter 4-Aromatic CompoundsNur Ayu Nadhirah Bt YahyaNo ratings yet
- Thermo 7e SM Chap01-1 ReduddcionDocument33 pagesThermo 7e SM Chap01-1 ReduddcionJonathanNo ratings yet
- Draft WPSDocument3 pagesDraft WPSWale OyeludeNo ratings yet
- Index 2015 Lithium-Process-ChemistryDocument8 pagesIndex 2015 Lithium-Process-ChemistryAlejandra RojasNo ratings yet
- AQA Physics Students Book AnswersDocument3 pagesAQA Physics Students Book AnswersT-Forty ArrowNo ratings yet
- CH 2Document3 pagesCH 2MD100% (1)
- BPG 01Document12 pagesBPG 01Ka Chun ChanNo ratings yet
- CementDocument5 pagesCementHemanth SaiNo ratings yet
- Surf Modif of PMMA Jet With H2ODocument6 pagesSurf Modif of PMMA Jet With H2OWilliams Marcel Caceres FerreiraNo ratings yet
- W2P Plate Filter PressDocument12 pagesW2P Plate Filter PressAngirishNo ratings yet
- Dossier Guanghua Casting PipeDocument58 pagesDossier Guanghua Casting PipeAlejandro C OlivaNo ratings yet
- Class 1CJ1S01 (Pip PN01CJ1S01 - 2007)Document4 pagesClass 1CJ1S01 (Pip PN01CJ1S01 - 2007)Zvonko BešlićNo ratings yet
- The Nature of Blue Brittleness of SteelDocument5 pagesThe Nature of Blue Brittleness of SteelAgustin Jose MarcuzziNo ratings yet
- Gas ProcessingDocument104 pagesGas ProcessingNicolae Marin100% (3)
- Pradhan Mantri Awas Yojana Housing For All (Urban) in AP StateDocument120 pagesPradhan Mantri Awas Yojana Housing For All (Urban) in AP StateKishore Nayak kNo ratings yet
- Chemistry Lab Manual 2Document21 pagesChemistry Lab Manual 2Nischay SanjayNo ratings yet
- Dow 2021 Australia Chemical RangeDocument4 pagesDow 2021 Australia Chemical Rangeneil smithiesNo ratings yet
- 4-Ductor: Insulated Conductor BarDocument13 pages4-Ductor: Insulated Conductor BarMitica TartareanuNo ratings yet