You might also like
- AJAB-2021!05!227 - Okay For GalleyDocument20 pagesAJAB-2021!05!227 - Okay For Galleyrania sheirNo ratings yet
- الجديد PDFDocument7 pagesالجديد PDFAwad AwadNo ratings yet
- International Journal of Food Microbiology 334 (2020) 108799Document11 pagesInternational Journal of Food Microbiology 334 (2020) 108799Tuğçe KanıkNo ratings yet
- In Vitro Anthelmintic Activity of The Crude Hydroalcoholic Extract of PiperDocument6 pagesIn Vitro Anthelmintic Activity of The Crude Hydroalcoholic Extract of PiperMarcial Fuentes EstradaNo ratings yet
- Insecticidal and Repellent Action of Pogostone Against Myzus Persicae (Hemiptera: Aphididae)Document5 pagesInsecticidal and Repellent Action of Pogostone Against Myzus Persicae (Hemiptera: Aphididae)Tarek DardouriNo ratings yet
- Mu Ma Et Al 2008Document6 pagesMu Ma Et Al 2008voskomilNo ratings yet
- MainDocument10 pagesMainKennVaneNo ratings yet
- Cárdenas-Manríquez Et Al. - 2016 - Environmental and Experimental BotanyDocument9 pagesCárdenas-Manríquez Et Al. - 2016 - Environmental and Experimental BotanyIsaac VegaNo ratings yet
- Insect Natural Enemies Can Develop Resistance To Insecticides in The Field Just As Their Hosts Ca1Document25 pagesInsect Natural Enemies Can Develop Resistance To Insecticides in The Field Just As Their Hosts Ca1Umair FaheemNo ratings yet
- tmpBA15 TMPDocument12 pagestmpBA15 TMPFrontiersNo ratings yet
- Tmpa936 TMPDocument12 pagesTmpa936 TMPFrontiersNo ratings yet
- Antifungal Activity of GuavaDocument11 pagesAntifungal Activity of GuavaFarij AbdurrohmanNo ratings yet
- Anthelmintic Activity of Acacia Oxyphylla Stem Bark Against Ascaridia GalliDocument7 pagesAnthelmintic Activity of Acacia Oxyphylla Stem Bark Against Ascaridia Gallifood heheNo ratings yet
- Benelli 2015Document12 pagesBenelli 2015Gustavo AndresNo ratings yet
- Fmicb 13 822689Document16 pagesFmicb 13 822689Ramanan ThirumoorthyNo ratings yet
- Thakur 2019Document11 pagesThakur 2019yasinNo ratings yet
- Residual Behavior and Dietary Risk Assessment of Albendazole As Fungicide in Citrus OrchardsDocument10 pagesResidual Behavior and Dietary Risk Assessment of Albendazole As Fungicide in Citrus OrchardsNat NewNo ratings yet
- Biofilm Inhibition and Anti Quorum Sensing Activity of Phytosynthesized Silver Nanoparticles Against The Nosocomial Pathogen Pseudomonas AeruginosaDocument17 pagesBiofilm Inhibition and Anti Quorum Sensing Activity of Phytosynthesized Silver Nanoparticles Against The Nosocomial Pathogen Pseudomonas AeruginosaReemaNo ratings yet
- Potential of Using Avonoids, Latex and Extracts From Calotropis Procera (Ait.) As Grain Protectants Against Two Coleopteran Pests of Stored RiceDocument8 pagesPotential of Using Avonoids, Latex and Extracts From Calotropis Procera (Ait.) As Grain Protectants Against Two Coleopteran Pests of Stored RiceglornumrNo ratings yet
- Martínez Et Al 2018Document10 pagesMartínez Et Al 2018Adrian Melgratti JobsonNo ratings yet
- Larvicidal Activity of Celosin A, A Novel Compound Isolated From Celosia Argentea LinnDocument6 pagesLarvicidal Activity of Celosin A, A Novel Compound Isolated From Celosia Argentea LinnInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- tmp330D TMPDocument18 pagestmp330D TMPFrontiersNo ratings yet
- Heliyon: Heeyoun Bunch, Jungeun Park, Hyeseung Choe, MD Munir Mosta Fiz, Jang-Eok Kim, Kyeong-Yeoll LeeDocument7 pagesHeliyon: Heeyoun Bunch, Jungeun Park, Hyeseung Choe, MD Munir Mosta Fiz, Jang-Eok Kim, Kyeong-Yeoll LeeMarioJuradoNo ratings yet
- Journal of Entomology and Zoology StudiesDocument11 pagesJournal of Entomology and Zoology StudiesAasha RanaNo ratings yet
- Hypericum Perforatum Plant Cells Reduce Agrobacterium ViabilityDocument8 pagesHypericum Perforatum Plant Cells Reduce Agrobacterium ViabilityFrontiersNo ratings yet
- Chemosphere: Jaqueline Bianchi, Thais Cristina Casimiro Fernandes, Maria Aparecida Marin-MoralesDocument9 pagesChemosphere: Jaqueline Bianchi, Thais Cristina Casimiro Fernandes, Maria Aparecida Marin-MoralesJose Franco NaviaNo ratings yet
- 2015 2 Lajmanovich y Col.Document8 pages2015 2 Lajmanovich y Col.Mariana CabagnaNo ratings yet
- Growth Performance, Caeca Microbial Population and Immune Response of Starter Broiler Chicks Fed Aqueous Extract of Balanites Aegyptiaca and Alchornea Cordifolia Stem Bark MixtureDocument9 pagesGrowth Performance, Caeca Microbial Population and Immune Response of Starter Broiler Chicks Fed Aqueous Extract of Balanites Aegyptiaca and Alchornea Cordifolia Stem Bark MixtureUIJRT United International Journal for Research & TechnologyNo ratings yet
- Aphis Craccivora (Koch) : Craccivora As One of The Main Pests, and ToDocument5 pagesAphis Craccivora (Koch) : Craccivora As One of The Main Pests, and ToAhmed Tarek SharafeldinNo ratings yet
- Cytological Effects of Blitox On Root Mitosis of Allium: Cepa LDocument7 pagesCytological Effects of Blitox On Root Mitosis of Allium: Cepa La lNo ratings yet
- Dang 2009Document13 pagesDang 2009EEDIEB Prof. Millton Marques CurvoNo ratings yet
- A Review Article: Anti-Quorum Sensing Agents As A Potential Replacement For Antibiotics in PhytobacteriologyDocument5 pagesA Review Article: Anti-Quorum Sensing Agents As A Potential Replacement For Antibiotics in PhytobacteriologyRajshree VermaNo ratings yet
- Singh 2010Document7 pagesSingh 2010Ismael Cervantes ChavezNo ratings yet
- Cell Growth Inhibitory Potential of Craterellus Cornucopioides (L.) Pers. Together With Antioxidant and Antimicrobial PropertiesDocument5 pagesCell Growth Inhibitory Potential of Craterellus Cornucopioides (L.) Pers. Together With Antioxidant and Antimicrobial Propertiesela.sofiaNo ratings yet
- Ca2 信号通路诱导的肠紧密连接蛋白受损 PDFDocument13 pagesCa2 信号通路诱导的肠紧密连接蛋白受损 PDFGaryNo ratings yet
- Wang Et Al 2012 - Toxicity of Neonics To Earthworms - Ecotox"0Saf AIPDocument7 pagesWang Et Al 2012 - Toxicity of Neonics To Earthworms - Ecotox"0Saf AIPFajana OyinlolaNo ratings yet
- Oliveira Et Al - South Africa Journal of BotanyDocument10 pagesOliveira Et Al - South Africa Journal of BotanyEDUARDO JOSÉ AZEVEDO CORREANo ratings yet
- Evaluation of Indigenous Farooq 2021Document7 pagesEvaluation of Indigenous Farooq 2021Ghulam SarwarNo ratings yet
- Saudi Journal of Biological SciencesDocument9 pagesSaudi Journal of Biological SciencesEvangelia SitanggangNo ratings yet
- Determination of Toxicity of Spinosad Against The Pulse Beetle, CallosobruchusDocument6 pagesDetermination of Toxicity of Spinosad Against The Pulse Beetle, CallosobruchustofazzalhNo ratings yet
- Subolesin Vaccine PDFDocument7 pagesSubolesin Vaccine PDFajithkumarkgNo ratings yet
- Poultrysci91 2148Document10 pagesPoultrysci91 2148Julio Enrique Ramirez HuancaNo ratings yet
- Antibacterial Mechanism of Lactic Acid On Physiological andDocument6 pagesAntibacterial Mechanism of Lactic Acid On Physiological andlox standardNo ratings yet
- DocumentDocument12 pagesDocumentariefNo ratings yet
- Biocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaDocument9 pagesBiocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaMaria BatoolNo ratings yet
- Agaricus Bisporus Fucogalactan Structural Characterization and Pharmacological ApproachesDocument8 pagesAgaricus Bisporus Fucogalactan Structural Characterization and Pharmacological ApproachesHuy NguyenNo ratings yet
- Benzofurano-Isatins: Search For Antimicrobial Agents: Arabian Journal of ChemistryDocument8 pagesBenzofurano-Isatins: Search For Antimicrobial Agents: Arabian Journal of Chemistrylucian_lovNo ratings yet
- Armando Et Al 2011Document10 pagesArmando Et Al 2011Adrian Melgratti JobsonNo ratings yet
- 2012-Bt Based Product Esterases and Phosphatase Larvae AedesDocument6 pages2012-Bt Based Product Esterases and Phosphatase Larvae AedesDwiNo ratings yet
- 1 s2.0 S138266892200165X MainDocument9 pages1 s2.0 S138266892200165X MainevelynNo ratings yet
- Mechanisms of Acaricide Resistance in The Cattle Tick Rhipicephalus (Boophilus) Microplus in SriLankaDocument5 pagesMechanisms of Acaricide Resistance in The Cattle Tick Rhipicephalus (Boophilus) Microplus in SriLankaAriadnaNo ratings yet
- Environmental Pollution: Manli Duan, Haichao Li, Jie Gu, Xiaxia Tuo, Wei Sun, Xun Qian, Xiaojuan WangDocument9 pagesEnvironmental Pollution: Manli Duan, Haichao Li, Jie Gu, Xiaxia Tuo, Wei Sun, Xun Qian, Xiaojuan WangGeorgiana-LuizaNo ratings yet
- Retrieve 19Document10 pagesRetrieve 19Nugroho WiratamaNo ratings yet
- Identification and Characterization Using Cytochrome p450Document11 pagesIdentification and Characterization Using Cytochrome p450jitenmechNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- Chemosphere: M. Govarthanan, Fuad Ameen, S. Kamala-Kannan, T. Selvankumar, A. Almansob, S.S. Alwakeel, Woong KimDocument7 pagesChemosphere: M. Govarthanan, Fuad Ameen, S. Kamala-Kannan, T. Selvankumar, A. Almansob, S.S. Alwakeel, Woong KimAwaliyah FajarNo ratings yet
- Talahib-Methanolic-Leaves-Not BslaDocument9 pagesTalahib-Methanolic-Leaves-Not BslaBunny BabeNo ratings yet
- MK Mycotoxins and PesticidesDocument47 pagesMK Mycotoxins and PesticidesrazadrpkNo ratings yet
- 82634-198862-1-PB Test Allium PDFDocument8 pages82634-198862-1-PB Test Allium PDFedoroftei2000No ratings yet
- Endophyte Biotechnology: Potential for Agriculture and PharmacologyFrom EverandEndophyte Biotechnology: Potential for Agriculture and PharmacologyAlexander SchoutenNo ratings yet
- 2020-Srikaya Seed Granule Different TemperatureDocument7 pages2020-Srikaya Seed Granule Different TemperatureDwiNo ratings yet
- 2014-Maybe Used-Ocotea CymbarumDocument10 pages2014-Maybe Used-Ocotea CymbarumDwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
- A Transcriptomic Survey of TheDocument20 pagesA Transcriptomic Survey of TheDwiNo ratings yet
- A Toxin-Binding Alkaline PhospDocument20 pagesA Toxin-Binding Alkaline PhospDwiNo ratings yet
- 2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDocument7 pages2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDwiNo ratings yet
- 2017-Justicia Adhatoda MidgutDocument11 pages2017-Justicia Adhatoda MidgutDwiNo ratings yet
- Action of Natural PhytosanitarDocument10 pagesAction of Natural PhytosanitarDwiNo ratings yet
- A Transformed Bacterium ExpresDocument16 pagesA Transformed Bacterium ExpresDwiNo ratings yet
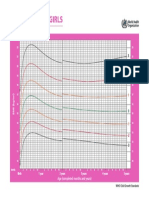
- CHT Bfa Girls Z 0 5Document1 pageCHT Bfa Girls Z 0 5DwiNo ratings yet
- Attachment 1706 631288cdc271dDocument5 pagesAttachment 1706 631288cdc271dDwiNo ratings yet
- 2012-Fungitoxic InsecticidalDocument18 pages2012-Fungitoxic InsecticidalDwiNo ratings yet
- CHT Bfa Boys Z 2 5Document1 pageCHT Bfa Boys Z 2 5DwiNo ratings yet
- 2012-CYP450 On EarthDocument12 pages2012-CYP450 On EarthDwiNo ratings yet
- Attachment 1706 631288cdc2e9fDocument4 pagesAttachment 1706 631288cdc2e9fDwiNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- 2012-Crimson Speckled MothDocument10 pages2012-Crimson Speckled MothDwiNo ratings yet
- 2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDocument682 pages2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDwiNo ratings yet
- 2012 Citrus Sinensis Musca DomesticaDocument8 pages2012 Citrus Sinensis Musca DomesticaDwiNo ratings yet
- Aedes Aegypti Bacillus ThuringiensisDocument9 pagesAedes Aegypti Bacillus ThuringiensisDwiNo ratings yet
- 2012-Castor Bean Leaf-Spodoptera LituraDocument7 pages2012-Castor Bean Leaf-Spodoptera LituraDwiNo ratings yet
- 2012-Calatropis GiganteaDocument14 pages2012-Calatropis GiganteaDwiNo ratings yet
- 2012-Bti SV2, Serratia Nematodipila SV6Document7 pages2012-Bti SV2, Serratia Nematodipila SV6DwiNo ratings yet
- 2012-Bti Residual TMOFDocument5 pages2012-Bti Residual TMOFDwiNo ratings yet
- 2012-Bti-Applied and Environmental MicrobiologyDocument6 pages2012-Bti-Applied and Environmental MicrobiologyDwiNo ratings yet
- Mechanical HandbookDocument48 pagesMechanical HandbookbenlandisNo ratings yet
- Spectrophotometric Method For Polyphenols Analysis Pre Validation and Application On Plantago L SpeciesDocument6 pagesSpectrophotometric Method For Polyphenols Analysis Pre Validation and Application On Plantago L SpeciesVan CarlosNo ratings yet
- Threads As Per IS 554: Pressure vs. Temperature Rating Chart For CPVC 4120 As Per ASTM D 2846Document3 pagesThreads As Per IS 554: Pressure vs. Temperature Rating Chart For CPVC 4120 As Per ASTM D 2846Asif S KhanNo ratings yet
- Ncert Booster Test SeriesDocument17 pagesNcert Booster Test SerieskailashNo ratings yet
- Design-II Samples Tutorial PDFDocument385 pagesDesign-II Samples Tutorial PDFHani Kirmani100% (1)
- EG-P For General Information - LC - EG-P - 20151001-002Document12 pagesEG-P For General Information - LC - EG-P - 20151001-002Lailatun Ni'mahNo ratings yet
- L6 PDFDocument17 pagesL6 PDFNilabha DasNo ratings yet
- Effi Calc F&TDocument8 pagesEffi Calc F&Tbronx_ldNo ratings yet
- Test Psictecnicos GratisDocument3 pagesTest Psictecnicos GratisLuis Eduardo CalelNo ratings yet
- Thermodynamics of Polymer Solutions: ΔG = Δh − T ΔsDocument29 pagesThermodynamics of Polymer Solutions: ΔG = Δh − T Δsdavid_tomy_1No ratings yet
- Ces 3Document5 pagesCes 3LegendaryNNo ratings yet
- Turning Gear An Operation Point of ViewDocument22 pagesTurning Gear An Operation Point of ViewAshwin Narayan100% (1)
- Lecture 19w Groundwater 1 Darcy PowerpointDocument19 pagesLecture 19w Groundwater 1 Darcy PowerpointAmir ShahzadNo ratings yet
- Reduction of Switching Losses in Igbt Power Modules (Dr. Thesis)Document86 pagesReduction of Switching Losses in Igbt Power Modules (Dr. Thesis)zeevko1249No ratings yet
- All1 58Document61 pagesAll1 58arisNo ratings yet
- Lec 08, Hydraulic Grade Line (HGL) and EnergyDocument8 pagesLec 08, Hydraulic Grade Line (HGL) and EnergyMahar Mamoon Rasheed100% (1)
- MCAT AAMC Content Outline - Science PDFDocument165 pagesMCAT AAMC Content Outline - Science PDFadfhNo ratings yet
- HjaimfgnhvDocument535 pagesHjaimfgnhvTrong DoanNo ratings yet
- IV Curve LabDocument14 pagesIV Curve Labapi-270103222No ratings yet
- Multiparameter Photometer With COD For Water and Wastewater: With Digital PH Electrode InputDocument6 pagesMultiparameter Photometer With COD For Water and Wastewater: With Digital PH Electrode InputXANPHOXYL KIBETNo ratings yet
- Impulse Forming by Vaporizing Foil ActuatorDocument35 pagesImpulse Forming by Vaporizing Foil ActuatorAnjithNo ratings yet
- How To Make Lye - 13 Steps - WikiHowDocument8 pagesHow To Make Lye - 13 Steps - WikiHowedalzurcNo ratings yet
- Student Exploration: PH Analysis: Lemon Juice Tastes Sour. When You Get Lemon Juice in Your Eye It BurnsDocument3 pagesStudent Exploration: PH Analysis: Lemon Juice Tastes Sour. When You Get Lemon Juice in Your Eye It BurnsHailey’s WoodNo ratings yet
- Characteristics of Sweet Potato Instant Cream Soup For Emergency FoodDocument8 pagesCharacteristics of Sweet Potato Instant Cream Soup For Emergency FoodRichal LaksmanaNo ratings yet
- HL BIO IB Extended EssayDocument39 pagesHL BIO IB Extended EssayRachel Tan Hui MinNo ratings yet
- MEM05052ADocument10 pagesMEM05052Arafiuddin100% (1)
- Module 7Document116 pagesModule 7adiNo ratings yet
- Use of Remote Indicators Is ForDocument11 pagesUse of Remote Indicators Is Forcaat_06No ratings yet
- RC Corbel Design (ACI318)Document2 pagesRC Corbel Design (ACI318)Emad KhanNo ratings yet
- Study On Scanning Micro-Arc Oxidation Technology Applied To 2024 Aluminum AlloyDocument6 pagesStudy On Scanning Micro-Arc Oxidation Technology Applied To 2024 Aluminum AlloyCicero Gomes de SouzaNo ratings yet