You might also like
- Anomalies in Geology. Physical, Chemical, BiologicalDocument344 pagesAnomalies in Geology. Physical, Chemical, BiologicalmancolNo ratings yet
- Applied Respiratory Physiology 2010 NUNNSDocument516 pagesApplied Respiratory Physiology 2010 NUNNSJaime Alberto Rodriguez100% (1)
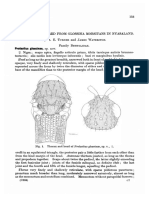
- Beetle: Beetle Beetles Are A Group of Insects That Form The Order Coleoptera, in TheDocument38 pagesBeetle: Beetle Beetles Are A Group of Insects That Form The Order Coleoptera, in Theenzo abrahamNo ratings yet
- CMI - FossilsDocument152 pagesCMI - Fossils7ett_60% (5)
- Proceedings of The 3rd IGCP 2013Document371 pagesProceedings of The 3rd IGCP 2013Aron SiccardiNo ratings yet
- Early dinosaur evolution and diversificationDocument8 pagesEarly dinosaur evolution and diversificationAjay SinghNo ratings yet
- DinosaurDocument23 pagesDinosaurD HernandezNo ratings yet
- The Mesozoic Era Age of DinosaursDocument196 pagesThe Mesozoic Era Age of DinosaursSANDRA BALLESTEROSNo ratings yet
- The Origin and Evolution of DinosaursDocument55 pagesThe Origin and Evolution of DinosaursTiago Savignon Cardoso MachadoNo ratings yet
- Downsized Dinosaurs: The Evolutionary Transition To Modern BirdsDocument9 pagesDownsized Dinosaurs: The Evolutionary Transition To Modern BirdsMyrthe de GoeijNo ratings yet
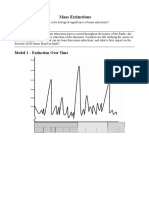
- Mass ExtinctionsDocument6 pagesMass ExtinctionsJacqueline50% (2)
- Paleontology GlossaryDocument24 pagesPaleontology GlossarycurlyfriezNo ratings yet
- Ancient Stem Reptiles: Cotylosaurs and PelycosaursDocument11 pagesAncient Stem Reptiles: Cotylosaurs and PelycosaursjeraldNo ratings yet
- Ichthyostega. Both Acanthostega and Ichthyostega Had More Than 5 Fingers On TheirDocument6 pagesIchthyostega. Both Acanthostega and Ichthyostega Had More Than 5 Fingers On TheirMarcosNo ratings yet
- Evolution of The Sirenia: An OutlineDocument10 pagesEvolution of The Sirenia: An Outlinelahsivlahsiv684No ratings yet
- ReptileDocument31 pagesReptileWira SentanuNo ratings yet
- Mesezoic EraDocument55 pagesMesezoic EraKingZhytt DrizNo ratings yet
- The History of Life in the Mesozoic EraDocument36 pagesThe History of Life in the Mesozoic EraSANDRA BALLESTEROSNo ratings yet
- Mammal: MammalsDocument13 pagesMammal: Mammalsdaitya99No ratings yet
- Flea Insecta Siphonaptera Family DiversityDocument15 pagesFlea Insecta Siphonaptera Family Diversitydhajaira.abNo ratings yet
- Fajfer 2012Document22 pagesFajfer 2012Monika FajferNo ratings yet
- Protozoa: Protozoa (From The Greek Words Proto, Meaning "First", and Zoa, Meaning "Animals") Is A GroupingDocument8 pagesProtozoa: Protozoa (From The Greek Words Proto, Meaning "First", and Zoa, Meaning "Animals") Is A Groupinggabrielluis08No ratings yet
- MammalDocument20 pagesMammalhandy wicaksanaNo ratings yet
- Trilobite Trilobites (Document29 pagesTrilobite Trilobites (enzo abrahamNo ratings yet
- Phylum ArthropodaDocument5 pagesPhylum ArthropodaMA. LYN CASIPENo ratings yet
- Arroyo-Cabrales Et Al (2007) - Overview of The Q Mammals From MXDocument14 pagesArroyo-Cabrales Et Al (2007) - Overview of The Q Mammals From MXArturo Palma RamírezNo ratings yet
- Oldest Predaceous Water Bugs Insecta Heteroptera Belostomatidae With Implications For Paleolimnology of The Triassic Cow Branch FormationDocument12 pagesOldest Predaceous Water Bugs Insecta Heteroptera Belostomatidae With Implications For Paleolimnology of The Triassic Cow Branch FormationDinaNo ratings yet
- Project 2 đề 1 The TimeDocument11 pagesProject 2 đề 1 The TimeKim Anh Đặng ThịNo ratings yet
- 360 Wake Annurev - Ecolsys 09Document20 pages360 Wake Annurev - Ecolsys 09bex9ggNo ratings yet
- Bernard ReviewerDocument2 pagesBernard ReviewerBernard BrowneNo ratings yet
- ArboniferousDocument1 pageArboniferousdes oroNo ratings yet
- Insectos Del Cenozoico en La ArgentinaDocument9 pagesInsectos Del Cenozoico en La ArgentinaDavid ArenasNo ratings yet
- DinosaurrrrsssDocument9 pagesDinosaurrrrsssjude albaoNo ratings yet
- Hylogeography of Inches and Parrows: Antonio Arnaiz-Villena, Pablo Gomez-Prieto and Valentin Ruiz-del-ValleDocument54 pagesHylogeography of Inches and Parrows: Antonio Arnaiz-Villena, Pablo Gomez-Prieto and Valentin Ruiz-del-ValleKakatuar KakatuaNo ratings yet
- 19bgi33c U3Document7 pages19bgi33c U3ArunNo ratings yet
- Abdala&Gaetano 2018 Late Triassic WorldDocument39 pagesAbdala&Gaetano 2018 Late Triassic WorldmioclaenusNo ratings yet
- A Review of The Nearctic Jumping SpidersDocument12 pagesA Review of The Nearctic Jumping SpidersGenaroNo ratings yet
- Dinosaurs: A Brief Background on Ruling ReptilesDocument1 pageDinosaurs: A Brief Background on Ruling ReptilesMatthymattNo ratings yet
- Theropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRDocument3 pagesTheropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRagomezgallardoNo ratings yet
- Artificial Keys To The Genera of Living Stalked CRDocument33 pagesArtificial Keys To The Genera of Living Stalked CRAurelia Izzatul AzkaNo ratings yet
- Origin of MammalsDocument26 pagesOrigin of Mammalsjai kumarNo ratings yet
- Phylogeny of The Terrestrial IsopodaDocument36 pagesPhylogeny of The Terrestrial IsopodaPedroNo ratings yet
- Kammerer 2020 Kongonaphon LagerpetidDocument5 pagesKammerer 2020 Kongonaphon Lagerpetidmauricio.garciaNo ratings yet
- 11Document25 pages11api-3713202No ratings yet
- The Early Mesosoic Era: Triassic (251-202 MYA) Jurassic (202-142 MYA)Document52 pagesThe Early Mesosoic Era: Triassic (251-202 MYA) Jurassic (202-142 MYA)andreea g.No ratings yet
- Echinoderm: Marine Animals With Radial SymmetryDocument1 pageEchinoderm: Marine Animals With Radial SymmetryhasoNo ratings yet
- Phylum ArthropodaDocument19 pagesPhylum ArthropodaOkta Novalia GNo ratings yet
- Coddington LeviSyst EvolSpiders91Document28 pagesCoddington LeviSyst EvolSpiders91LUIS MIGUEL OSORIO VANEGASNo ratings yet
- Coddington LeviSyst EvolSpiders91Document28 pagesCoddington LeviSyst EvolSpiders91Luis OsorioNo ratings yet
- GastropodsDocument12 pagesGastropodsLeoregine RodriguezNo ratings yet
- Mass Extinction of BirdsDocument5 pagesMass Extinction of BirdsnomadNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- DinosaurDocument31 pagesDinosaurAngielou SardualNo ratings yet
- Sishe Salsabila Luqyana Practicum ReportDocument27 pagesSishe Salsabila Luqyana Practicum ReportSalsabila LuqyanaNo ratings yet
- Reptiles Are: Hylonomus CasineriaDocument1 pageReptiles Are: Hylonomus CasineriaAlesha BishunNo ratings yet
- Group 6 - ScienceDocument11 pagesGroup 6 - ScienceYuuchaNo ratings yet
- MesozoikumDocument44 pagesMesozoikumGregoryNo ratings yet
- The Phylogeny of Hexapoda Arthropoda and PDFDocument15 pagesThe Phylogeny of Hexapoda Arthropoda and PDFResita ReiitaNo ratings yet
- Wiki Loves MonumentsDocument7 pagesWiki Loves MonumentsKvb SenabsolzurcNo ratings yet
- Phylogeny and Biogeography of Asian Pit VipersDocument32 pagesPhylogeny and Biogeography of Asian Pit VipersHaydee ParasNo ratings yet
- Mammal: Jump To Navigationjump To SearchDocument15 pagesMammal: Jump To Navigationjump To SearchYeyen M. EvoraNo ratings yet
- Early evolution and diversification of dinosaursDocument1 pageEarly evolution and diversification of dinosaursELYAHWEH kongsi KebenaranNo ratings yet
- Palaeogeography, Palaeoclimatology, Palaeoecology: Alexandra A.E. Van Der Geer, George A. Lyras, Rebekka VolmerDocument12 pagesPalaeogeography, Palaeoclimatology, Palaeoecology: Alexandra A.E. Van Der Geer, George A. Lyras, Rebekka Volmer-Dwiki Mulya-No ratings yet
- Scannella & Horner 2010Document13 pagesScannella & Horner 2010elenabarreto885No ratings yet
- Triassic Life on Land: The Great TransitionFrom EverandTriassic Life on Land: The Great TransitionRating: 4 out of 5 stars4/5 (1)
- Thrips (Thysanoptera, Insecta) From The Rovno Amber, Eocene of UkraineDocument6 pagesThrips (Thysanoptera, Insecta) From The Rovno Amber, Eocene of UkraineJean-Bernard HuchetNo ratings yet
- Shodotova Pyralid Buryatia 2008Document10 pagesShodotova Pyralid Buryatia 2008Jean-Bernard HuchetNo ratings yet
- Shattuck 2009 Formicidae Gen NDocument7 pagesShattuck 2009 Formicidae Gen NJean-Bernard HuchetNo ratings yet
- Shaw 2003 Megalyridae Rev CryptalyraDocument4 pagesShaw 2003 Megalyridae Rev CryptalyraJean-Bernard HuchetNo ratings yet
- Sha Current Research Cretaceous China Lake 2007Document9 pagesSha Current Research Cretaceous China Lake 2007Jean-Bernard HuchetNo ratings yet
- Abrahamson Et Al 1998 Cynipidae Gall Inducing QuercusDocument7 pagesAbrahamson Et Al 1998 Cynipidae Gall Inducing QuercusJean-Bernard HuchetNo ratings yet
- A New Genus and Species of Fungus-Gnats (My Cetophilidae) - F. RDocument3 pagesA New Genus and Species of Fungus-Gnats (My Cetophilidae) - F. RJean-Bernard HuchetNo ratings yet
- SHARKEY - 2005 - Braconidae Agathirsia SP N AgathidinaeDocument5 pagesSHARKEY - 2005 - Braconidae Agathirsia SP N AgathidinaeJean-Bernard HuchetNo ratings yet
- Revision of blister beetle generaDocument14 pagesRevision of blister beetle generaJean-Bernard HuchetNo ratings yet
- Effects of Larval Density on Flesh Fly Life CyclesDocument9 pagesEffects of Larval Density on Flesh Fly Life CyclesJean-Bernard HuchetNo ratings yet
- Turner Waterston 1916 Bethylidae Rev ProlaeliusDocument3 pagesTurner Waterston 1916 Bethylidae Rev ProlaeliusJean-Bernard HuchetNo ratings yet
- James 1966 Stratiomyidae Pachygastrinae Gen NDocument5 pagesJames 1966 Stratiomyidae Pachygastrinae Gen NJean-Bernard HuchetNo ratings yet
- Ponomarenko Ren 2010 Cupedidae Notocupes Jurassic ChinaDocument3 pagesPonomarenko Ren 2010 Cupedidae Notocupes Jurassic ChinaJean-Bernard HuchetNo ratings yet
- Oswald 1990 Hemerobiidae Nomerobius Rev GenDocument13 pagesOswald 1990 Hemerobiidae Nomerobius Rev GenJean-Bernard HuchetNo ratings yet
- Oswald 1989 Psychopsidae Nom Gen RevDocument9 pagesOswald 1989 Psychopsidae Nom Gen RevJean-Bernard HuchetNo ratings yet
- Ahmad Ghani 1974 Encyrtidae Gen NDocument4 pagesAhmad Ghani 1974 Encyrtidae Gen NJean-Bernard HuchetNo ratings yet
- Amat, 2010 - Notes On Necrophagous Flies Associated To Fish Carrion in Colombian AmazonDocument4 pagesAmat, 2010 - Notes On Necrophagous Flies Associated To Fish Carrion in Colombian AmazonJean-Bernard HuchetNo ratings yet
- Zehner Et Al., 2004 - Genetic Identification of Forensically Important Flesh FliesDocument3 pagesZehner Et Al., 2004 - Genetic Identification of Forensically Important Flesh FliesJean-Bernard HuchetNo ratings yet
- Li Et Al. 2019 - ElectrorubesopsinaeDocument10 pagesLi Et Al. 2019 - ElectrorubesopsinaeJean-Bernard HuchetNo ratings yet
- Mafart Et Al., 2004 - 3D Imaging Hominids StudiesDocument7 pagesMafart Et Al., 2004 - 3D Imaging Hominids StudiesJean-Bernard HuchetNo ratings yet
- Peck, 2009 - Beetle Species Diversity in The Lesser Antilles IslandsDocument8 pagesPeck, 2009 - Beetle Species Diversity in The Lesser Antilles IslandsJean-Bernard HuchetNo ratings yet
- A Review of The Genus Glaresis ErichsonDocument17 pagesA Review of The Genus Glaresis ErichsonJean-Bernard HuchetNo ratings yet
- Ames Et Al., 2006 - The Use of Mitochondrial Cytochrome Oxidase I Gene (COI) To Differentiate C. Vicina Et C. VomitoriaDocument4 pagesAmes Et Al., 2006 - The Use of Mitochondrial Cytochrome Oxidase I Gene (COI) To Differentiate C. Vicina Et C. VomitoriaJean-Bernard HuchetNo ratings yet
- Cockerell 1917 F Insects Florissant ColoradoDocument4 pagesCockerell 1917 F Insects Florissant ColoradoJean-Bernard HuchetNo ratings yet
- Wells & Sperling, 2001 - DNA Based Identification of Forensically Important ChrysomyinaeDocument6 pagesWells & Sperling, 2001 - DNA Based Identification of Forensically Important ChrysomyinaeJean-Bernard HuchetNo ratings yet
- DiZinno Et Al., 2002 - Mitochondrial DNA Sequencing of Beetle Larvae (Omosita) Recovered From Human BoneDocument3 pagesDiZinno Et Al., 2002 - Mitochondrial DNA Sequencing of Beetle Larvae (Omosita) Recovered From Human BoneJean-Bernard HuchetNo ratings yet
- Harvey Et Al., 2008 - A Global Study of Forensically Significant CalliphoridsDocument12 pagesHarvey Et Al., 2008 - A Global Study of Forensically Significant CalliphoridsJean-Bernard HuchetNo ratings yet
- Huchet & Greenberg 2010 Flies, Mochicas and Burial Practices, A Case Study From Huaca de La Luna, PeruDocument11 pagesHuchet & Greenberg 2010 Flies, Mochicas and Burial Practices, A Case Study From Huaca de La Luna, PeruJean-Bernard HuchetNo ratings yet
- Bernath & Kunz, 1981 - Structure and Dynamics of Arthropod Communities in Bat Guano Deposits in BuildingsDocument11 pagesBernath & Kunz, 1981 - Structure and Dynamics of Arthropod Communities in Bat Guano Deposits in BuildingsJean-Bernard HuchetNo ratings yet
- Reading Comprehension ExerciseDocument21 pagesReading Comprehension ExercisehidampradigaraNo ratings yet
- Geology of Salt RangeDocument18 pagesGeology of Salt RangeAdil KhanNo ratings yet
- Amonites Turonianos de TexasDocument454 pagesAmonites Turonianos de TexasKevin Andres Gonzalez AvellanedaNo ratings yet
- Indian Stratigraphic Boundary ProblemsDocument6 pagesIndian Stratigraphic Boundary ProblemsArpit UpadhyayNo ratings yet
- Chap 22Document42 pagesChap 22Parham ElahiNo ratings yet
- Diversity of Life Through Time: Diversification of Life Patterns - Marine, Nonmarine, Vertebrates, PlantsDocument7 pagesDiversity of Life Through Time: Diversification of Life Patterns - Marine, Nonmarine, Vertebrates, PlantsnareshNo ratings yet
- 24 Mass Extinctions-SDocument16 pages24 Mass Extinctions-SRomilPatelNo ratings yet
- Vision IAS Prelims Test 18 Solutions 2022Document36 pagesVision IAS Prelims Test 18 Solutions 2022ulaganathanNo ratings yet
- Synthesizing Sources on Healthy Eating HabitsDocument9 pagesSynthesizing Sources on Healthy Eating HabitsKAREN ONGCOL JUMALONNo ratings yet
- Chapter 8: Geologic Time: Thinking About Time The History of (Relative) Time Geologic Time Numerical Time Rates of ChangeDocument45 pagesChapter 8: Geologic Time: Thinking About Time The History of (Relative) Time Geologic Time Numerical Time Rates of ChangeKen AguilaNo ratings yet
- Osmosis Fall 2022Document15 pagesOsmosis Fall 2022Lily DicksonNo ratings yet
- Transient Metazoan Reefs in The Aftermath of The End-Permian Mass ExtinctionDocument5 pagesTransient Metazoan Reefs in The Aftermath of The End-Permian Mass ExtinctiontriassicpaperNo ratings yet
- The Chanthaburi Terrane of SoutheasternDocument17 pagesThe Chanthaburi Terrane of SoutheasternKyipyarNo ratings yet
- 11948355Document72 pages11948355Emin KarabasicNo ratings yet
- Subhendu Bardhan and Devapriya ChattopadhyayDocument28 pagesSubhendu Bardhan and Devapriya ChattopadhyayAlly MoralesNo ratings yet
- 3 s2.0 B9780124095489121165 MainDocument11 pages3 s2.0 B9780124095489121165 MainVictor VelascoNo ratings yet
- Salt RangeDocument35 pagesSalt RangeMuhammad Waqas89% (18)
- Salida 14Document3 pagesSalida 14Cristian Camilo Rincón Hernández100% (1)
- Master Series Mock Cat 1 PDFDocument90 pagesMaster Series Mock Cat 1 PDFShubham KabraNo ratings yet
- The Lilliput Effect in The Aftermath of The End-Permian Extinction EventDocument13 pagesThe Lilliput Effect in The Aftermath of The End-Permian Extinction Eventapi-3713202No ratings yet
- Zimmer Tangled Bank Chapter 10 With NoticeDocument30 pagesZimmer Tangled Bank Chapter 10 With Noticecorneliohd34No ratings yet
- Machine Learning Identifies Ecological Selectivity Patterns Across The End Permian Mass Extinction PDFDocument15 pagesMachine Learning Identifies Ecological Selectivity Patterns Across The End Permian Mass Extinction PDFPooja ReddyNo ratings yet
- The 5 Major Mass Extinctions that Shaped Life on EarthDocument6 pagesThe 5 Major Mass Extinctions that Shaped Life on EarthHarun RashidNo ratings yet
- Biodiversity PDFDocument24 pagesBiodiversity PDFstaldNo ratings yet