You might also like
- Salmonellae Interactions With Host Process NATUREDocument15 pagesSalmonellae Interactions With Host Process NATUREDiegoNo ratings yet
- Velgeetal 2012Document17 pagesVelgeetal 2012shaznay delacruzNo ratings yet
- Salmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008Document8 pagesSalmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008noordin MukasaNo ratings yet
- Toxocara Canis Molecular Basis of Immune RecognitiDocument10 pagesToxocara Canis Molecular Basis of Immune RecognitiBarbie GuerreroNo ratings yet
- Helmintos TisularesDocument8 pagesHelmintos TisularesValentina Arenas RNo ratings yet
- Articulo 3 - SalmonellaDocument16 pagesArticulo 3 - SalmonellaJoiver Alean FlórezNo ratings yet
- Helmintos NematodosDocument12 pagesHelmintos NematodosHANNIANo ratings yet
- PatogenesisDocument28 pagesPatogenesisOka WidyaningsihNo ratings yet
- Salmonella Enterica Serovar Ty-Phimurium: Biology and Diseases of Ruminants: Sheep, Goats, and CattleDocument10 pagesSalmonella Enterica Serovar Ty-Phimurium: Biology and Diseases of Ruminants: Sheep, Goats, and CattleMegitsu NavaNo ratings yet
- Dissemination of Invasive Salmonella Via Bacterial-Induced Extrusion of Mucosal EpitheliaDocument6 pagesDissemination of Invasive Salmonella Via Bacterial-Induced Extrusion of Mucosal Epitheliashaznay delacruzNo ratings yet
- Paper Seminario 01Document10 pagesPaper Seminario 01Hans MenaresNo ratings yet
- Salmonella Enterica Serovars Typhimurium and Typhi As Model OrganismsDocument14 pagesSalmonella Enterica Serovars Typhimurium and Typhi As Model OrganismsShaznay Dela CruzNo ratings yet
- 2A - Sarigumba-Scenario For Think BreakDocument7 pages2A - Sarigumba-Scenario For Think BreakSean SarigumbaNo ratings yet
- Immunology of the Toxoplasma gondii ParasiteDocument17 pagesImmunology of the Toxoplasma gondii ParasiteVic PastorNo ratings yet
- Genomic Analysis of An Outbreak Isolate of Salmonella Entericaserovarenteritidis ST11 Fromcayar, SenegalDocument6 pagesGenomic Analysis of An Outbreak Isolate of Salmonella Entericaserovarenteritidis ST11 Fromcayar, SenegalIJAR JOURNALNo ratings yet
- PHD Thesis SalmonellaDocument4 pagesPHD Thesis Salmonellafqzaoxjef100% (2)
- Salmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerDocument14 pagesSalmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerShaznay Dela CruzNo ratings yet
- Colera 2Document13 pagesColera 2Cindy GarciaNo ratings yet
- International Journal For ParasitologyDocument10 pagesInternational Journal For ParasitologyGabriel AvilaNo ratings yet
- Jurnal Patogenesis AmoebiasisDocument12 pagesJurnal Patogenesis AmoebiasisAngga NuralamNo ratings yet
- Accepted Manuscript: Molecular & Biochemical ParasitologyDocument25 pagesAccepted Manuscript: Molecular & Biochemical ParasitologyColo VoltaNo ratings yet
- 2004 Macrophages, Pathology and Parasite Persistence in Experimental Visceral LeishmaniasisDocument7 pages2004 Macrophages, Pathology and Parasite Persistence in Experimental Visceral LeishmaniasismclimacoNo ratings yet
- Molecular Mechanisms Used by Salmonella To Evade The Immune SystemDocument35 pagesMolecular Mechanisms Used by Salmonella To Evade The Immune SystemANDREA FERNANDA MUÑOZ NARVAEZNo ratings yet
- Journal SchistosomiasisDocument11 pagesJournal SchistosomiasisCelissa Mauriz HilarioNo ratings yet
- Ch24 - Host Resistance To Gastrointestinal1Document12 pagesCh24 - Host Resistance To Gastrointestinal1g8ksdvpt9rNo ratings yet
- Hussain 2021Document13 pagesHussain 2021Amín MoraNo ratings yet
- لغات التواصل الطفيلي (1) Document24 pagesلغات التواصل الطفيلي (1) Houda BouachaNo ratings yet
- Literature Review On SalmonellaDocument6 pagesLiterature Review On Salmonellac5r9j6zj100% (1)
- The Biochemistry of Malaria: Yongyuth Yuthavong and Prapon WilairatDocument6 pagesThe Biochemistry of Malaria: Yongyuth Yuthavong and Prapon Wilairataliyu belloNo ratings yet
- The Genome of Salmonella enterica Serovar TyphiDocument5 pagesThe Genome of Salmonella enterica Serovar TyphiguemadiNo ratings yet
- Fasciola Hepatica-Derived Molecules As Potential InmunomodulatorsDocument7 pagesFasciola Hepatica-Derived Molecules As Potential Inmunomodulatorsvalitajs28No ratings yet
- Kvir 14 2158656Document22 pagesKvir 14 2158656Robert StryjakNo ratings yet
- 575 Full PDFDocument12 pages575 Full PDFandualemNo ratings yet
- Uso de Vacunas de ADN vs. Salmonella EntericaDocument10 pagesUso de Vacunas de ADN vs. Salmonella EntericaalferezhassanNo ratings yet
- Salmonella, The Host and Disease: A Brief ReviewDocument8 pagesSalmonella, The Host and Disease: A Brief ReviewPedro Albán MNo ratings yet
- Do GargaloDocument9 pagesDo GargaloGustavo FelpeNo ratings yet
- Control of Human ToxoplasmosisDocument27 pagesControl of Human ToxoplasmosisCarissima djaraNo ratings yet
- Fcimb 11 662766Document10 pagesFcimb 11 662766Gustavo AndresNo ratings yet
- Salmonella Infection of Gallbladder Epithelial Cells Drives Local Inflammation and Injury in A Model of Acute Typhoid FeverDocument38 pagesSalmonella Infection of Gallbladder Epithelial Cells Drives Local Inflammation and Injury in A Model of Acute Typhoid FeverAgussalimNo ratings yet
- Epithelial Inflammasomes Defend Against Salmonella Gut InfectionDocument9 pagesEpithelial Inflammasomes Defend Against Salmonella Gut InfectionShaznay Dela CruzNo ratings yet
- Genome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andDocument7 pagesGenome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andArsh McBarsh MalhotraNo ratings yet
- Quorum Sensing in African Trypanosomes: SciencedirectDocument6 pagesQuorum Sensing in African Trypanosomes: SciencedirectNithin SakthiNo ratings yet
- Pathogenesis in Tuberculosis: Transcriptomic Approaches To Unraveling Virulence Mechanisms and Finding New Drug TargetsDocument23 pagesPathogenesis in Tuberculosis: Transcriptomic Approaches To Unraveling Virulence Mechanisms and Finding New Drug Targetskinta translateNo ratings yet
- ExosomesinfectiousdiseaseDocument10 pagesExosomesinfectiousdiseaseTaniaNo ratings yet
- 145 WqeDocument8 pages145 WqeKatia RamónNo ratings yet
- Kvir 13 2074130Document33 pagesKvir 13 2074130Robert StryjakNo ratings yet
- Genomic Virulence Genes Profile Analysis of Salmonella Enterica Isolates From Animal and Human in China From 2004 To 2019Document9 pagesGenomic Virulence Genes Profile Analysis of Salmonella Enterica Isolates From Animal and Human in China From 2004 To 2019LuisNo ratings yet
- Club 04Document13 pagesClub 04Muñoz Cerón Smiht YhoissNo ratings yet
- Omaña-Molina Et Al., Type 2 DM BALB-c Mice Are More Susceptible To GAEDocument10 pagesOmaña-Molina Et Al., Type 2 DM BALB-c Mice Are More Susceptible To GAEIsmael CastelanNo ratings yet
- 07 For Shell 541554Document14 pages07 For Shell 541554frankyNo ratings yet
- Review Vacunas Toxo 2019 PDFDocument15 pagesReview Vacunas Toxo 2019 PDFVic PastorNo ratings yet
- Macropinocytosis and SARS-CoV-2 Cell EntryDocument12 pagesMacropinocytosis and SARS-CoV-2 Cell EntryKarla SegovianoNo ratings yet
- Cei 12674Document10 pagesCei 12674mclimacoNo ratings yet
- HHS Public Access: The Lytic Cycle of Toxoplasma Gondii: 15 Years LaterDocument27 pagesHHS Public Access: The Lytic Cycle of Toxoplasma Gondii: 15 Years Laterjuan pablo zuluaga montoyaNo ratings yet
- Effector ReviewDocument26 pagesEffector ReviewSumit MukherjeeNo ratings yet
- Parasita No Intestino Do MosquitoDocument8 pagesParasita No Intestino Do MosquitoGustavo FelpeNo ratings yet
- Animal Models of Salmonella Infections e PDFDocument10 pagesAnimal Models of Salmonella Infections e PDFaminzaghiNo ratings yet
- Mammalian Cell Culture: Lisa Sandell and Daisuke SakaiDocument32 pagesMammalian Cell Culture: Lisa Sandell and Daisuke SakaiJitendra Singh VermaNo ratings yet
- Rna ActivityDocument3 pagesRna ActivityJassy GonzalesNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- Pramoolsinsap 1998Document6 pagesPramoolsinsap 1998Shaznay Dela CruzNo ratings yet
- Reviews The Role of Intestinal Endotoxin in Liver Injury: A Long and Evolving HistoryDocument7 pagesReviews The Role of Intestinal Endotoxin in Liver Injury: A Long and Evolving HistoryShaznay Dela CruzNo ratings yet
- A Model For Bacterial Pathogenesis: AlmonellaDocument18 pagesA Model For Bacterial Pathogenesis: AlmonellaShaznay Dela CruzNo ratings yet
- Endotoxin, Reticuloendothelial Function, And: Liver InjuryDocument8 pagesEndotoxin, Reticuloendothelial Function, And: Liver InjuryShaznay Dela CruzNo ratings yet
- Bitar 1985Document15 pagesBitar 1985Shaznay Dela CruzNo ratings yet
- Hepatic Dysfunction in Paediatric Typhoidal Salmonellosis: Annals of Tropical PaediatricsDocument4 pagesHepatic Dysfunction in Paediatric Typhoidal Salmonellosis: Annals of Tropical PaediatricsShaznay Dela CruzNo ratings yet
- Multidrug-Resistant Bovine Salmonellosis PredisposDocument6 pagesMultidrug-Resistant Bovine Salmonellosis PredisposShaznay Dela CruzNo ratings yet
- Interactions Between Bacteria and Bile Salts in THDocument13 pagesInteractions Between Bacteria and Bile Salts in THShaznay Dela CruzNo ratings yet
- Salmonella Enterica Serovars Typhimurium and Typhi As Model OrganismsDocument14 pagesSalmonella Enterica Serovars Typhimurium and Typhi As Model OrganismsShaznay Dela CruzNo ratings yet
- Epithelial Inflammasomes Defend Against Salmonella Gut InfectionDocument9 pagesEpithelial Inflammasomes Defend Against Salmonella Gut InfectionShaznay Dela CruzNo ratings yet
- Ribet 2015Document11 pagesRibet 2015Shaznay Dela CruzNo ratings yet
- Intestinal Mucosa: Through The Lens of The OrganismDocument10 pagesIntestinal Mucosa: Through The Lens of The OrganismShaznay Dela CruzNo ratings yet
- Salmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerDocument14 pagesSalmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerShaznay Dela CruzNo ratings yet
- Zhou 2001Document6 pagesZhou 2001Shaznay Dela CruzNo ratings yet
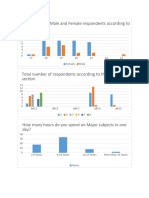
- Gender and age survey results of respondentsDocument3 pagesGender and age survey results of respondentsShaznay Dela CruzNo ratings yet
- Philippine Disaster Management WorksheetsDocument9 pagesPhilippine Disaster Management WorksheetsJen Dela CruzNo ratings yet
- A Detailed Lesson Plan in Biology (Grade 8) I. Objectives: Daily RoutineDocument7 pagesA Detailed Lesson Plan in Biology (Grade 8) I. Objectives: Daily RoutineShaznay Dela CruzNo ratings yet
- Gender and age survey results of respondentsDocument3 pagesGender and age survey results of respondentsShaznay Dela CruzNo ratings yet
- BIOCHEMDocument6 pagesBIOCHEMShaznay Dela CruzNo ratings yet
- Special Stains in Histopath: Cryptococcus NeoformansDocument3 pagesSpecial Stains in Histopath: Cryptococcus NeoformansLUZVIMINDA GORDONo ratings yet
- Bacteria Collection - NCTC 10400 Bacillus Subtilis Subsp SpizizeniiDocument2 pagesBacteria Collection - NCTC 10400 Bacillus Subtilis Subsp Spizizeniijair gonzales puenteNo ratings yet
- Dwnload Full Fundamentals of Anatomy and Physiology 8th Edition Martini Test Bank PDFDocument36 pagesDwnload Full Fundamentals of Anatomy and Physiology 8th Edition Martini Test Bank PDFwaterycobstone.yg9zl100% (11)
- Plant and Animal Organ Systems FunctionsDocument18 pagesPlant and Animal Organ Systems FunctionsHannah CastroNo ratings yet
- Mtap - Special Histopath TechniquesDocument21 pagesMtap - Special Histopath TechniquesK-idol LiveNo ratings yet
- 3D Electron Microscopy TechniquesDocument1 page3D Electron Microscopy TechniquesMuraleetharan_BNo ratings yet
- Unknown Lab Report-2Document16 pagesUnknown Lab Report-2api-508768911No ratings yet
- DLP 2nd Quarter Week 1 2 Page 1Document6 pagesDLP 2nd Quarter Week 1 2 Page 1Junriel Arig BonachitaNo ratings yet
- GenBio1 Module1Document10 pagesGenBio1 Module1Alexander DontonNo ratings yet
- PLANT and ANIMAL CELLSDocument16 pagesPLANT and ANIMAL CELLSjoan ruby bautista100% (1)
- Andrefsky-A Brief Introduction To The Lithic AnalysisDocument10 pagesAndrefsky-A Brief Introduction To The Lithic AnalysisggarfuNo ratings yet
- BIO104E-Laboratory Activity. (The Cell)Document5 pagesBIO104E-Laboratory Activity. (The Cell)Stephen AzaresNo ratings yet
- CytoskeletonDocument65 pagesCytoskeletonapi-19916399No ratings yet
- ANA205 Muscular Tissue: Lecture Notes by Dr. Taiwo-OlaDocument31 pagesANA205 Muscular Tissue: Lecture Notes by Dr. Taiwo-OlaIbrahim BabatundeNo ratings yet
- Prokaryotic Cell Structure and FunctionsDocument39 pagesProkaryotic Cell Structure and Functionsabida waryamNo ratings yet
- AP Module4 Trans DiazDocument18 pagesAP Module4 Trans DiazJohn Rafael DiazNo ratings yet
- FomeDocument8 pagesFomemirelamanteamirelaNo ratings yet
- 1) BioMEMSDocument150 pages1) BioMEMShjunior89No ratings yet
- The MicroscopeDocument6 pagesThe MicroscopeFat PatNo ratings yet
- General Biology1 For Grade 11 StudentsDocument39 pagesGeneral Biology1 For Grade 11 StudentsInero PHNo ratings yet
- OMAX MD827S30L-X 879TYR Microscope User ManualDocument19 pagesOMAX MD827S30L-X 879TYR Microscope User ManualmelfamaniaNo ratings yet
- Q2 LONG TEST 1 MicroscopeDocument5 pagesQ2 LONG TEST 1 MicroscopeCastolo Bayucot JvjcNo ratings yet
- Advances in Microscopy Techniques: Review ArticleDocument9 pagesAdvances in Microscopy Techniques: Review Articlekajol22fulNo ratings yet
- Cell Structure TarsiaDocument2 pagesCell Structure TarsiaNurul'IzzahNo ratings yet
- TEORI DASAR BIOLOGI SELDocument36 pagesTEORI DASAR BIOLOGI SELanon_862299899No ratings yet
- Surface ChemistryDocument137 pagesSurface ChemistryMohammed86% (7)
- Immunofluorescence - Types, Techniques, and Limitations - New York Microscope CompanyDocument6 pagesImmunofluorescence - Types, Techniques, and Limitations - New York Microscope CompanyMuhammed SabdatNo ratings yet
- 10.1007/978 3 319 01360 2Document333 pages10.1007/978 3 319 01360 2Bheim Llona100% (1)
- Biology Quiz on Cells, Cell Theory and OrganismsDocument16 pagesBiology Quiz on Cells, Cell Theory and Organismsgcjar vllnNo ratings yet
- Em CCD910012Document2 pagesEm CCD910012mythNo ratings yet