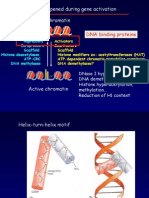
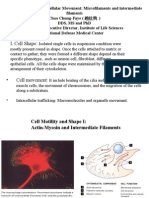
You might also like
- Fatty Acid-Derived Signals in Plants: Hans WeberDocument8 pagesFatty Acid-Derived Signals in Plants: Hans WeberQuenguan FranklinNo ratings yet
- Mira Et al-2009-FEMS Yeast ResearchDocument15 pagesMira Et al-2009-FEMS Yeast ResearchRita JanelaNo ratings yet
- Recent Developments in Carbapenems: ReviewDocument16 pagesRecent Developments in Carbapenems: ReviewFrancielleNo ratings yet
- Ahn Et Al 2010 Electrophilic Tuning of The Chemoprotective Natural Product SulforaphaneDocument6 pagesAhn Et Al 2010 Electrophilic Tuning of The Chemoprotective Natural Product Sulforaphane1766636545No ratings yet
- Chow 2009Document12 pagesChow 2009Nadia SaiNo ratings yet
- Paper 3Document5 pagesPaper 3AntoniaNo ratings yet
- Mechanistic Studies of The Inactivation of TEM-1 and P99 by NXL104, A Novel Non - Lactam - Lactamase InhibitorDocument7 pagesMechanistic Studies of The Inactivation of TEM-1 and P99 by NXL104, A Novel Non - Lactam - Lactamase InhibitorSALMA MAGALÍ DE LA ROSA POSADASNo ratings yet
- Indian Journal 1Document4 pagesIndian Journal 1rajiNo ratings yet
- Membrane-Binding Mechanism of Clostridium Perfringens Alpha-ToxinDocument8 pagesMembrane-Binding Mechanism of Clostridium Perfringens Alpha-ToxinTanvir AhmedNo ratings yet
- Arber and LinnDocument7 pagesArber and Linnshubham20No ratings yet
- 2008 - Purification, crystallization and preliminary X-ray diffraction analysis of a cystathionine β-synthase domain-containing protein, CDCP2, from Arabidopsis thalianaDocument3 pages2008 - Purification, crystallization and preliminary X-ray diffraction analysis of a cystathionine β-synthase domain-containing protein, CDCP2, from Arabidopsis thalianado258No ratings yet
- AssstDocument5 pagesAssstDiegoNo ratings yet
- Chlorophyll Degradation in A Chlamydomonas Reinhardtii Mutant An Accumulation of Pyropheophorbide A by Anaerobiosis, Doi Et Al., 2001Document6 pagesChlorophyll Degradation in A Chlamydomonas Reinhardtii Mutant An Accumulation of Pyropheophorbide A by Anaerobiosis, Doi Et Al., 2001deryhermawanNo ratings yet
- Calcium Spirulan, An Inhibitor of Enveloped Virus Replication, From A Blue-GreenDocument5 pagesCalcium Spirulan, An Inhibitor of Enveloped Virus Replication, From A Blue-GreenDiana Valeria Pimentel AguilarNo ratings yet
- Properties of Copper-Free Pig Kidney Amine Oxidase: Role of Topa QuinoneDocument8 pagesProperties of Copper-Free Pig Kidney Amine Oxidase: Role of Topa QuinoneGizwa AiichaicchapratiwieNo ratings yet
- Distinct Function of Pseudomonas Aeruginosa TypeDocument5 pagesDistinct Function of Pseudomonas Aeruginosa TypejohanNo ratings yet
- One-Step Measurement of Firefly Luciferase Activity in YeastDocument5 pagesOne-Step Measurement of Firefly Luciferase Activity in Yeastapi-272992171No ratings yet
- Capitulo 27Document47 pagesCapitulo 27Ana Paola SámanoNo ratings yet
- Phosphorylation of The Human MicroRNA Generating C PDFDocument11 pagesPhosphorylation of The Human MicroRNA Generating C PDFcgilgasNo ratings yet
- RNA Silencing: 90th International Titisee ConferenceDocument9 pagesRNA Silencing: 90th International Titisee ConferenceViệt KhoaNo ratings yet
- 3.5 7.3 7.4 Transcription Translation Answers 1Document8 pages3.5 7.3 7.4 Transcription Translation Answers 1Zuza PodleśnaNo ratings yet
- Cell. Mol. Life Sci. (2009) 66:3697-3710 DOI 10.1007/s00018-009-0103-6Document14 pagesCell. Mol. Life Sci. (2009) 66:3697-3710 DOI 10.1007/s00018-009-0103-6mlbrenciagliaNo ratings yet
- 10 05 2006 Lifescience 2Document60 pages10 05 2006 Lifescience 2api-3696530No ratings yet
- Biochemical and Molecular Characterization of The Venom From The Cuban Scorpion Rhopalurus JunceusDocument10 pagesBiochemical and Molecular Characterization of The Venom From The Cuban Scorpion Rhopalurus JunceusCapitulina AmbrioletaNo ratings yet
- Phosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalDocument12 pagesPhosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalNini FarmNo ratings yet
- Operon SourceDocument4 pagesOperon Sourcemustafa aliNo ratings yet
- Article: Jasmonate Perception by Inositol-Phosphate-Potentiated COI1-JAZ Co-ReceptorDocument8 pagesArticle: Jasmonate Perception by Inositol-Phosphate-Potentiated COI1-JAZ Co-ReceptorMariz SintahaNo ratings yet
- TMP 36 CBDocument6 pagesTMP 36 CBFrontiersNo ratings yet
- Function, Structure and Therapeutic Potential of Complement C5a ReceptorsDocument20 pagesFunction, Structure and Therapeutic Potential of Complement C5a ReceptorsFrontiersNo ratings yet
- Wound Responsive Cis Element in The 5 Upstream Region of Cucumber Ascorbate Oxidase GeneDocument8 pagesWound Responsive Cis Element in The 5 Upstream Region of Cucumber Ascorbate Oxidase GenenurNo ratings yet
- Ribosome-Targeting Antibiotics and Mechanisms of Bacterial ResistanceDocument14 pagesRibosome-Targeting Antibiotics and Mechanisms of Bacterial ResistanceColeguillasNo ratings yet
- Turkmen 2011Document6 pagesTurkmen 2011Arefin ShahriarNo ratings yet
- Synthesis and in Vitro Antiprotozoal Activities of 5-Phenyliminobenzo (A) Phenoxazine DerivativesDocument4 pagesSynthesis and in Vitro Antiprotozoal Activities of 5-Phenyliminobenzo (A) Phenoxazine Derivativesfaz7No ratings yet
- Pi Is 1097276515000027Document13 pagesPi Is 1097276515000027Yeny Benedetti RosalesNo ratings yet
- Plant Physiol Chapter 23 - Abscisic Acid - A Seed Maturation and Antistress SignalDocument20 pagesPlant Physiol Chapter 23 - Abscisic Acid - A Seed Maturation and Antistress SignalBrunoDeminicisNo ratings yet
- Ref 04Document11 pagesRef 04truong.nguyen.19.12.92No ratings yet
- Streptococcus Faecalis: Membrane Adenosine Triphosphatase FromDocument8 pagesStreptococcus Faecalis: Membrane Adenosine Triphosphatase FromSinta 2712No ratings yet
- Escherichia ColiDocument8 pagesEscherichia ColiKanishk PandeyNo ratings yet
- Membrane Binding of The Bacterial Signal Recognition Particle Receptor Involves Two Distinct Binding SitesDocument10 pagesMembrane Binding of The Bacterial Signal Recognition Particle Receptor Involves Two Distinct Binding SitesDeeptha MadhavanNo ratings yet
- Lecture6-Central DogmaDocument79 pagesLecture6-Central DogmaAlberto Laborte Jr.No ratings yet
- 8 Sigmas2003Document7 pages8 Sigmas2003AsmaNo ratings yet
- 12 08 2006 MCB Cytoskeleton and Movement II 1Document129 pages12 08 2006 MCB Cytoskeleton and Movement II 1api-3696530No ratings yet
- Cold Adaptation Proteins: Characterization, The Heat-Labile From Antarctic PsychrophileDocument6 pagesCold Adaptation Proteins: Characterization, The Heat-Labile From Antarctic PsychrophileSelfi YantiNo ratings yet
- Bioorganic & Medicinal Chemistry LettersDocument5 pagesBioorganic & Medicinal Chemistry LettersRhysnaChemsNo ratings yet
- Genetic Characterisation of The Botulinurn Toxin Complex ofDocument8 pagesGenetic Characterisation of The Botulinurn Toxin Complex ofnurhasanahNo ratings yet
- Use of Phosphonyl Carbanions in The Synthesis of Anti-Inflammatory Active Phosphorus-Containing Fused Heterocycles and Relevance PhosphonateDocument8 pagesUse of Phosphonyl Carbanions in The Synthesis of Anti-Inflammatory Active Phosphorus-Containing Fused Heterocycles and Relevance PhosphonateSelenia ReyesNo ratings yet
- A Flavin-Based Extracellular Electron Transfer Mechanism in Diverse Gram-Positive Bacteria REFERENCEDocument19 pagesA Flavin-Based Extracellular Electron Transfer Mechanism in Diverse Gram-Positive Bacteria REFERENCENvo SpektarNo ratings yet
- Proteina de Elongacion eEf1ADocument5 pagesProteina de Elongacion eEf1ABertha Calderón-LimónNo ratings yet
- Aurora & MitosisDocument11 pagesAurora & MitosisJubairNo ratings yet
- Photosynthesis: Topic 1Document9 pagesPhotosynthesis: Topic 1rahmansulayman56No ratings yet
- Viper Venom Botox The Molecular Origin ADocument4 pagesViper Venom Botox The Molecular Origin AAmmar SansourNo ratings yet
- Role of Sigma FactorDocument29 pagesRole of Sigma Factorsandhya namadaraNo ratings yet
- Quorum-Sensing-Dependent Regulation of Biosynthesis of The Polyketide Antibiotic Mupirocin in Pseudomonas Fluorescens NCIMB 10586Document13 pagesQuorum-Sensing-Dependent Regulation of Biosynthesis of The Polyketide Antibiotic Mupirocin in Pseudomonas Fluorescens NCIMB 10586Venny ChenNo ratings yet
- TMP 2 FEDocument2 pagesTMP 2 FEFrontiersNo ratings yet
- Hkale Biology - 2 The CytologyDocument11 pagesHkale Biology - 2 The Cytologynovember flushNo ratings yet
- FDADocument7 pagesFDASachithNo ratings yet
- Bioorganic & Medicinal ChemistryDocument7 pagesBioorganic & Medicinal ChemistryRodrigo Sebastian Arancibia GonzalezNo ratings yet
- Cro Regulatory ProteinDocument7 pagesCro Regulatory ProteinvijayNo ratings yet
- A.I. Yaropolou Analytica Chimica Acta 308 (1995) 137-144Document8 pagesA.I. Yaropolou Analytica Chimica Acta 308 (1995) 137-144CLAUDIA SICHELNo ratings yet
- Tetrahedron Reports on Organic Chemistry: Volume 4.31-40From EverandTetrahedron Reports on Organic Chemistry: Volume 4.31-40Derek BartonNo ratings yet
- 2 +Aisy+RahmaniaDocument9 pages2 +Aisy+RahmaniaKaka TaeNo ratings yet
- Microrev00035 0201Document19 pagesMicrorev00035 0201Kaka TaeNo ratings yet
- Jenis Lalat Buah Bactrocera Spp. (Diptera: Tephritidae) Yang Menyerang Pertanaman Jeruk Di Kabupaten Gianyar Dan BangliDocument9 pagesJenis Lalat Buah Bactrocera Spp. (Diptera: Tephritidae) Yang Menyerang Pertanaman Jeruk Di Kabupaten Gianyar Dan BangliKaka TaeNo ratings yet
- HohoDocument2 pagesHohoKaka TaeNo ratings yet
- 1246-Article Text-2089-2-10-20161228Document10 pages1246-Article Text-2089-2-10-20161228Kaka TaeNo ratings yet
- Fitri Andalus Handayani & Mohamad Ichsana Nur 1 Implementasi Good Governance Di IndonesiaDocument11 pagesFitri Andalus Handayani & Mohamad Ichsana Nur 1 Implementasi Good Governance Di IndonesiaMeizy AnggunNo ratings yet
- It All Started With That NightDocument1 pageIt All Started With That NightKaka TaeNo ratings yet
- List of Famous Cities On River Banks in The WorldDocument2 pagesList of Famous Cities On River Banks in The WorldDiptangshu DeNo ratings yet
- Movie Review of THORDocument8 pagesMovie Review of THORSiva LetchumiNo ratings yet
- Fall 20122Document98 pagesFall 20122DobarCovek67% (3)
- DTS Nozzles R3Document2 pagesDTS Nozzles R3meilia teknikNo ratings yet
- Titus Selection of DiffuserDocument14 pagesTitus Selection of DiffuserhanyassawyNo ratings yet
- José Guadalupe PosadaDocument19 pagesJosé Guadalupe PosadaJudy Baca100% (1)
- GCG Damri SurabayaDocument11 pagesGCG Damri SurabayaEndang SusilawatiNo ratings yet
- p1632 eDocument4 pagesp1632 ejohn saenzNo ratings yet
- Panel Hospital List IGIDocument6 pagesPanel Hospital List IGIAbdul RahmanNo ratings yet
- Epidemiological Triad of HIV/AIDS: AgentDocument8 pagesEpidemiological Triad of HIV/AIDS: AgentRakib HossainNo ratings yet
- Paso de Blas Lying in Clinic For NewDocument5 pagesPaso de Blas Lying in Clinic For NewNaheed Dean MustafaNo ratings yet
- Ijrpr2741 Study On Investor Perception Towards Stock Market InvestmentDocument19 pagesIjrpr2741 Study On Investor Perception Towards Stock Market InvestmentAbhay RanaNo ratings yet
- Auburn Bsci ThesisDocument5 pagesAuburn Bsci Thesisafksaplhfowdff100% (1)
- Marketing Measurement Done RightDocument16 pagesMarketing Measurement Done RightWasim Ullah0% (1)
- JHS Integrated Science Preamble, Jan 2012 - FinalDocument15 pagesJHS Integrated Science Preamble, Jan 2012 - Finalfrank adamsNo ratings yet
- Maryam Ejaz Sec-A Marketing Assignment (CHP #15)Document3 pagesMaryam Ejaz Sec-A Marketing Assignment (CHP #15)MaryamNo ratings yet
- Subsea Pipeline Job DescriptionDocument2 pagesSubsea Pipeline Job DescriptionVijay_DamamNo ratings yet
- Episode Transcript: Episode 34 - Chinese Han Lacquer CupDocument2 pagesEpisode Transcript: Episode 34 - Chinese Han Lacquer CupParvathy SubramanianNo ratings yet
- Stitch2421 1 Kit PDFDocument8 pagesStitch2421 1 Kit PDFJoshua Robertson100% (1)
- Ken Holt 06 The Secret of Hangman's InnDocument216 pagesKen Holt 06 The Secret of Hangman's InnPastPresentFuture100% (2)
- Case Study Presentation - CGDocument37 pagesCase Study Presentation - CGapi-290866384No ratings yet
- Asme Bladder Accumulator DatasheetDocument3 pagesAsme Bladder Accumulator DatasheetSamad A BakarNo ratings yet
- THE Ketofeed Diet Book v2Document43 pagesTHE Ketofeed Diet Book v2jacosta12100% (1)
- Engineering Mathematics Questions and AnswersDocument9 pagesEngineering Mathematics Questions and AnswersZaky Muzaffar100% (1)
- FO - Course Transition Info - Acknowledgement - SIT50422 - Dip Hospitality Management - V1Document2 pagesFO - Course Transition Info - Acknowledgement - SIT50422 - Dip Hospitality Management - V1nitesh.kc06No ratings yet
- APS PresentationDocument32 pagesAPS PresentationRozack Ya ZhackNo ratings yet
- Wind LoadingDocument18 pagesWind LoadingStephen Ogalo100% (1)
- CS1 Entity Level Controls SolutionsDocument16 pagesCS1 Entity Level Controls SolutionsPakistan Breaking News100% (6)
- EHR StandardsIndia - August 2013-32630521Document54 pagesEHR StandardsIndia - August 2013-32630521kartiksinhNo ratings yet