You might also like
- Avian BiologyFrom EverandAvian BiologyDonald FarnerRating: 5 out of 5 stars5/5 (1)
- Insect MorphologyDocument25 pagesInsect MorphologySree Naveena100% (1)
- cá sấuDocument57 pagescá sấuHạt Vừng LépNo ratings yet
- Elife 43828Document19 pagesElife 43828jaypasco79No ratings yet
- Bones Paternes CaracaraDocument9 pagesBones Paternes Caracarasilviahfestamagica1No ratings yet
- 09 The Evolution of Flight in Bats A Novel HypothesisDocument14 pages09 The Evolution of Flight in Bats A Novel Hypothesisberenice simonNo ratings yet
- Amphioxus As A Model To Study The Evolution of DevDocument15 pagesAmphioxus As A Model To Study The Evolution of DevHeloísaNo ratings yet
- Anna Rufa Ruth Blasco AvesDocument13 pagesAnna Rufa Ruth Blasco AvesNegan DeanNo ratings yet
- Early Morphological Specialization For Insect-Spider Associations in Mesozoic LacewingsDocument24 pagesEarly Morphological Specialization For Insect-Spider Associations in Mesozoic Lacewingsstar warsNo ratings yet
- ArtiDocument13 pagesArtiSebàstian LopezNo ratings yet
- Introduction To The Skeleton of HummingbirdsDocument95 pagesIntroduction To The Skeleton of HummingbirdsCamilo Barrios CardozoNo ratings yet
- A New Genus and Species of Scorpionfly (Mecoptera) From Baltic Amber, With An Unusually Developed Postnotal OrganDocument6 pagesA New Genus and Species of Scorpionfly (Mecoptera) From Baltic Amber, With An Unusually Developed Postnotal Organagnieszka.soszynska.majNo ratings yet
- Ciclostomi FosiliDocument5 pagesCiclostomi FosiliAnca MihalcescuNo ratings yet
- Liu Et Al. 2017 - Flight Aerodynamics in Enantiornithines - Information From A New Chinese Early Cretaceous BirdDocument18 pagesLiu Et Al. 2017 - Flight Aerodynamics in Enantiornithines - Information From A New Chinese Early Cretaceous BirdDavidCernyNo ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- Phyl Ogen y of Phylum ChordataDocument10 pagesPhyl Ogen y of Phylum Chordatamirandajealyn28No ratings yet
- 10 1016@j Geobios 2020 09 005Document36 pages10 1016@j Geobios 2020 09 005DirtyBut WholeNo ratings yet
- Form, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaDocument10 pagesForm, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaSixto Gutiérrez SaavedraNo ratings yet
- Zusi2013 - Introduction To The Skeleton of Hummingbirds PDFDocument95 pagesZusi2013 - Introduction To The Skeleton of Hummingbirds PDFCamilo Barrios CardozoNo ratings yet
- Montalvo2011 PDFDocument8 pagesMontalvo2011 PDFPedro Dante RuizNo ratings yet
- Amphibian Larvae Morphology GuideDocument16 pagesAmphibian Larvae Morphology GuideMorgana OliveiraNo ratings yet
- American Ornithologists' UnionDocument15 pagesAmerican Ornithologists' UnionntoungwaNo ratings yet
- Zoology Paper 1 Topic Wise Upsc Questions: Syllabus Non-Chordata and ChordataDocument16 pagesZoology Paper 1 Topic Wise Upsc Questions: Syllabus Non-Chordata and ChordataThe Nish100% (2)
- Evidence Rats Out Bat EvolutionDocument3 pagesEvidence Rats Out Bat Evolutioneustoque2668No ratings yet
- UrochordatevolutionDocument15 pagesUrochordatevolutionRodrigo CarralNo ratings yet
- Fossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsDocument38 pagesFossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsRenz Junyll ApigoNo ratings yet
- Bambiraptor and DeinonychusDocument10 pagesBambiraptor and DeinonychusFelipe MesVasNo ratings yet
- Letter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaDocument4 pagesLetter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaThiago MarinhoNo ratings yet
- The Anatomical Record - 2021 - L Pez Aguirre - Variation in Cross Sectional Shape and Biomechanical Properties of The BatDocument16 pagesThe Anatomical Record - 2021 - L Pez Aguirre - Variation in Cross Sectional Shape and Biomechanical Properties of The BatLEIAH MARIE JOY RIONo ratings yet
- Journal of Anatomy - 2019 - Echeverr ADocument13 pagesJournal of Anatomy - 2019 - Echeverr Apjames199705No ratings yet
- Mayr Et Al-2016-PalaeontologyDocument11 pagesMayr Et Al-2016-PalaeontologyJorge BarNo ratings yet
- Evolution of The JawDocument13 pagesEvolution of The JawCraig MawsonNo ratings yet
- Philippine Birds of Prey: Interrelations Among Habitat, Morphology and BehaviorDocument14 pagesPhilippine Birds of Prey: Interrelations Among Habitat, Morphology and Behaviorgerman guazaNo ratings yet
- ใบงานปลาDocument83 pagesใบงานปลาpijitra senakhamNo ratings yet
- Pnas 2109210118Document12 pagesPnas 2109210118NedNo ratings yet
- Higher-Order Phylogeny of Modern Birds Comparative Anatomy BradleyDocument95 pagesHigher-Order Phylogeny of Modern Birds Comparative Anatomy BradleyMateusNo ratings yet
- The Evolution of Vertebrate Diversity: Chapter ObjectivesDocument14 pagesThe Evolution of Vertebrate Diversity: Chapter Objectivesirene9tan9ailianNo ratings yet
- The Scorpionandits VenomDocument67 pagesThe Scorpionandits VenomGabriel FloreaNo ratings yet
- Assignment_2.2Document2 pagesAssignment_2.2gkanamarlapudiNo ratings yet
- Origin and Evolution of Sound Production in SerrasalmidaeDocument12 pagesOrigin and Evolution of Sound Production in SerrasalmidaeJhon Philip BagaNo ratings yet
- Byers1988aDocument10 pagesByers1988aNouman AliNo ratings yet
- 2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDocument16 pages2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDredeltoNo ratings yet
- The Evolution of Early Vertebrate Photoreceptors: ReviewDocument16 pagesThe Evolution of Early Vertebrate Photoreceptors: ReviewJ.D. NobleNo ratings yet
- UntitledDocument104 pagesUntitledRain NakagoNo ratings yet
- Buasso Et Al. 2006 Predation On Snakes of Argentina, Effects of Coloration and Ring Pattern On Coral and False Coral SnakesDocument7 pagesBuasso Et Al. 2006 Predation On Snakes of Argentina, Effects of Coloration and Ring Pattern On Coral and False Coral SnakesAlejandra Niño ReyesNo ratings yet
- Yang HR 2021 Cretaceous Enigmatic Insect Magicivena SPPDocument11 pagesYang HR 2021 Cretaceous Enigmatic Insect Magicivena SPP杨楠No ratings yet
- Fenton, M (2012)Document11 pagesFenton, M (2012)lina maaria lopezNo ratings yet
- DamianDocument2 pagesDamianSTEPHANIE LOUISE DAMIANNo ratings yet
- The Tubercles On Humpback Whales' FlippersDocument11 pagesThe Tubercles On Humpback Whales' FlippersRNo ratings yet
- Specialized Adaptations For Springtail Predation in Mesozoic BeetlesDocument7 pagesSpecialized Adaptations For Springtail Predation in Mesozoic BeetlesArina DamayantiNo ratings yet
- Slooh CharracDocument30 pagesSlooh CharracTD TVNo ratings yet
- (Chiappe & Witmer) - Mesozoic Birds - 1st EditionDocument533 pages(Chiappe & Witmer) - Mesozoic Birds - 1st EditionCamila Pineda100% (1)
- Comparative Anatomy, Embryology & Vestigial StructuresDocument11 pagesComparative Anatomy, Embryology & Vestigial StructuresDaniel BerryNo ratings yet
- AfrocersiskenyensisGOK PublicationDocument8 pagesAfrocersiskenyensisGOK PublicationAyan DuttaNo ratings yet
- How To Make A Bird Skull, Major Transition in The Ecolution of The Avian Cranium, Paedomorphosis, and The Beak As A Surrogate HandDocument15 pagesHow To Make A Bird Skull, Major Transition in The Ecolution of The Avian Cranium, Paedomorphosis, and The Beak As A Surrogate HandMyrthe de GoeijNo ratings yet
- ITN Birds Are DinosDocument62 pagesITN Birds Are DinosAnonymous na2AlIuGNo ratings yet
- Brill: BRILL Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To BehaviourDocument47 pagesBrill: BRILL Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Behaviourapi-283358168No ratings yet
- Arthropod Structure & Development: Fabian B Aumler, Stanislav N. Gorb, Sebastian BüsseDocument12 pagesArthropod Structure & Development: Fabian B Aumler, Stanislav N. Gorb, Sebastian BüsseSuhar TatiNo ratings yet
- Bat Flight: Aerodynamics, Kinematics and Flight Morphology: Anders Hedenstro ̈ M and L. Christoffer JohanssonDocument11 pagesBat Flight: Aerodynamics, Kinematics and Flight Morphology: Anders Hedenstro ̈ M and L. Christoffer JohanssonMondoskisNo ratings yet
- DePalma Et Al 2019 A Seismically Induced Onshore Surge Deposit at The KG Boundary North DakotaDocument10 pagesDePalma Et Al 2019 A Seismically Induced Onshore Surge Deposit at The KG Boundary North DakotaMaría ChavarríaNo ratings yet
- Bertin Et Al Current Perspectives On Tooth Implantation Attahment and Replacement in AmniotaDocument20 pagesBertin Et Al Current Perspectives On Tooth Implantation Attahment and Replacement in AmniotaMaría ChavarríaNo ratings yet
- Agnolit Et Al 2019 Pre-Gabi Biotic Connections Between The Americas and Alternative Model To Explain The Less-Splendid Isolation of South AmreicaDocument16 pagesAgnolit Et Al 2019 Pre-Gabi Biotic Connections Between The Americas and Alternative Model To Explain The Less-Splendid Isolation of South AmreicaMaría ChavarríaNo ratings yet
- Cernansly 2020 The First Dibamidae From MongoliaDocument18 pagesCernansly 2020 The First Dibamidae From MongoliaMaría ChavarríaNo ratings yet
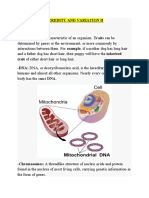
- Heredity and Variation IIDocument18 pagesHeredity and Variation IIsmbdy tbhhhNo ratings yet
- Autism and Vaccination (Final)Document8 pagesAutism and Vaccination (Final)Rajan JattNo ratings yet
- PedsCases CF Note (SP Edit)Document1 pagePedsCases CF Note (SP Edit)Mehtab AlamNo ratings yet
- The Medtechs EmeDocument3 pagesThe Medtechs EmeKathlene Anne MENDOZANo ratings yet
- Cell SignalingDocument38 pagesCell SignalingShyrene Mamanao GumbanNo ratings yet
- Bacterial InfectionsDocument6 pagesBacterial InfectionsErika Noreen Dela RosaNo ratings yet
- Benedicta G. Capunong: Peterpaul - Nacua@deped - Gov.phDocument11 pagesBenedicta G. Capunong: Peterpaul - Nacua@deped - Gov.phMichael MillanesNo ratings yet
- Nihms 1679935Document33 pagesNihms 1679935Rebeca Tapia RsNo ratings yet
- 1 s2.0 S2773111123000098 MainDocument21 pages1 s2.0 S2773111123000098 MainRavindu AriyarathneNo ratings yet
- Eoc Review PacketDocument12 pagesEoc Review Packetapi-293001217No ratings yet
- Zuhad by Rida Saleem CompleteDocument38 pagesZuhad by Rida Saleem CompleteAfrah NajeebNo ratings yet
- 10th Science QN Sep-20Document11 pages10th Science QN Sep-20santhoshsanthosh20202021No ratings yet
- May 2016 MS TZ0Document13 pagesMay 2016 MS TZ0Eda ŞafakNo ratings yet
- Biology 400 MCQs in English - (WWW - GkNotesPDF.com)Document26 pagesBiology 400 MCQs in English - (WWW - GkNotesPDF.com)jeetendra sahuNo ratings yet
- Day-17-PEDIGREE ANALYSIS KEYDocument2 pagesDay-17-PEDIGREE ANALYSIS KEYalishNo ratings yet
- De Novo Assembly of Agave Sisalana Transcriptome in Response To Drought StressDocument14 pagesDe Novo Assembly of Agave Sisalana Transcriptome in Response To Drought StressGregorio SamsaNo ratings yet
- General Zoology: For MidwivesDocument40 pagesGeneral Zoology: For MidwivesDanica Rose Daza MacahiloNo ratings yet
- DNA Properties UV AbsorptionDocument6 pagesDNA Properties UV AbsorptionSamreen SiddiquiNo ratings yet
- Obradovic, Et Al ('21) - Leptin and Obesity - Role and Clinical Implication (Fie)Document14 pagesObradovic, Et Al ('21) - Leptin and Obesity - Role and Clinical Implication (Fie)aida-69No ratings yet
- Wastewater surveillance of pathogens can inform public health responsesDocument4 pagesWastewater surveillance of pathogens can inform public health responsesMiriam LeiNo ratings yet
- Lycopene Food Sources - Potential Role in Human Health and Antioxidant EffectsDocument153 pagesLycopene Food Sources - Potential Role in Human Health and Antioxidant EffectsCoțovanu IulianNo ratings yet
- The Snapping Turtle Chelydra SerpentinaDocument2 pagesThe Snapping Turtle Chelydra Serpentinaapi-615380330No ratings yet
- REVIEWER - Biological PsychologyDocument59 pagesREVIEWER - Biological PsychologyMendoza Bernalyn DCNo ratings yet
- Chapter 13Document56 pagesChapter 13Hậu VũNo ratings yet
- Non-Mendelian Inheritance: Chapter OutlineDocument20 pagesNon-Mendelian Inheritance: Chapter OutlineEMANUEL DAVID COLMENARES LARANo ratings yet
- Visions of The Future WorksheetDocument2 pagesVisions of The Future WorksheetKevin Joseph PerfectoNo ratings yet
- Introductory Biochemistry: CHEM 160Document31 pagesIntroductory Biochemistry: CHEM 160Gianna Kristen MirandaNo ratings yet
- Context CluesDocument7 pagesContext CluesasherNo ratings yet
- Altered Immune ResponseDocument11 pagesAltered Immune ResponseRittika SinhaNo ratings yet
- Human Anatomy Physiology 8th Edition Marieb Test BankDocument34 pagesHuman Anatomy Physiology 8th Edition Marieb Test BankKevinHarrisonnfixq100% (14)