You might also like
- Angelova 2013 Review Models of Obesity and Metabolic SyndromeDocument8 pagesAngelova 2013 Review Models of Obesity and Metabolic SyndromePaul SimononNo ratings yet
- Considerations When Choosing High Fat High Fructose and H - 2021 - Current DevDocument10 pagesConsiderations When Choosing High Fat High Fructose and H - 2021 - Current DevFelipe MoitaNo ratings yet
- LandgrafDocument10 pagesLandgrafcicifaraditasuliNo ratings yet
- Amini Et Al, 2010Document7 pagesAmini Et Al, 2010Jesana LopesNo ratings yet
- Resveratrol Improves Health and Survival of Mice On A High-Calorie DietDocument6 pagesResveratrol Improves Health and Survival of Mice On A High-Calorie Dietapi-119817014No ratings yet
- Meal Frequency and Timing in Health and DiseaseDocument8 pagesMeal Frequency and Timing in Health and DiseaseaesocidNo ratings yet
- Jci37385 PDFDocument13 pagesJci37385 PDFNur Ahmad HabibiNo ratings yet
- Perubahan Berat Badan Tikus Sprague DawleyDocument9 pagesPerubahan Berat Badan Tikus Sprague DawleyFatma AdlifaNo ratings yet
- 1 s2.0 S2314808X16300033 MainDocument8 pages1 s2.0 S2314808X16300033 MainnisceNo ratings yet
- Sciencedirect: Caloric Dose-Responsive Genes in Blood Cells Differentiate The Metabolic Status of Obese MenDocument10 pagesSciencedirect: Caloric Dose-Responsive Genes in Blood Cells Differentiate The Metabolic Status of Obese MenWa Ode Umrawati LatifNo ratings yet
- Delade 2018Document9 pagesDelade 2018probo asmoroNo ratings yet
- 2020 UPFs Obesity Metaanalysis IJODocument12 pages2020 UPFs Obesity Metaanalysis IJOKelly Viviana Gaitan OyolaNo ratings yet
- Spirulina Reduces Diet-Induced Obesity Through Downregulation of Lipogenic Genes Expression inDocument10 pagesSpirulina Reduces Diet-Induced Obesity Through Downregulation of Lipogenic Genes Expression inMtra. Mayra N. Elizalde VillarrealNo ratings yet
- 1 s2.0 S0031938416309179 Main PDFDocument7 pages1 s2.0 S0031938416309179 Main PDFvharsyaNo ratings yet
- Diet-Induced Metabolic SyndromeDocument8 pagesDiet-Induced Metabolic SyndromeMatuar Hi UmarNo ratings yet
- 1 s2.0 S0955286317303868 MainDocument14 pages1 s2.0 S0955286317303868 MainAdib FraNo ratings yet
- FullDocument25 pagesFullAndrés Felipe Zapata MurielNo ratings yet
- Aliment Pharmacol Ther - 2020 - Logan - Analysis of 61 Exclusive Enteral Nutrition Formulas Used in The Management ofDocument13 pagesAliment Pharmacol Ther - 2020 - Logan - Analysis of 61 Exclusive Enteral Nutrition Formulas Used in The Management ofRohma YuliaNo ratings yet
- Anim Models and Exp Med - 2023 - LiDocument10 pagesAnim Models and Exp Med - 2023 - LiLevi MagalhãesNo ratings yet
- Efectos A Nivel Cardiovascular y CerebralDocument14 pagesEfectos A Nivel Cardiovascular y CerebralCRISTOFER EDUARDO GONZALEZ IRARRAZABALNo ratings yet
- Which Hyperglycemic Model of Zebrafish (Danio Rerio) Suites My Type 2 Diabetes Mellitus Research? A Scoring System For Available MethodsDocument15 pagesWhich Hyperglycemic Model of Zebrafish (Danio Rerio) Suites My Type 2 Diabetes Mellitus Research? A Scoring System For Available MethodsVero ArellanoNo ratings yet
- 2 ESPEN-ESPGHAN-ECFS Guidelines On Nutrition Care For Infants Children and Adults With Cystic FibrosisDocument21 pages2 ESPEN-ESPGHAN-ECFS Guidelines On Nutrition Care For Infants Children and Adults With Cystic FibrosisAmany SalamaNo ratings yet
- Schreyer 1998Document8 pagesSchreyer 1998Jocilene Dantas Torres NascimentoNo ratings yet
- 2 A Canela Atenua A Adiposidade e Afeta A ExpressãoDocument11 pages2 A Canela Atenua A Adiposidade e Afeta A Expressãoguto hdNo ratings yet
- Wong 2017Document8 pagesWong 2017diasrashmi653No ratings yet
- Proceedings 02 00573 v3Document186 pagesProceedings 02 00573 v3André SartorelliNo ratings yet
- 1 s2.0 S1110663815000592 MainDocument7 pages1 s2.0 S1110663815000592 Mainaminm23kNo ratings yet
- Comparison With Ancestral Diets Suggests Dense Acellular Carbohydrates Promote An Inflammatory Microbiota and May Be The Primary Dietary Cause of PDFDocument16 pagesComparison With Ancestral Diets Suggests Dense Acellular Carbohydrates Promote An Inflammatory Microbiota and May Be The Primary Dietary Cause of PDFFernando MaffeisNo ratings yet
- Docosahexaenoic Acid Administered in The Acute Phase Protects Nutritional Status of Septic Neonates - Nutrition 2006Document7 pagesDocosahexaenoic Acid Administered in The Acute Phase Protects Nutritional Status of Septic Neonates - Nutrition 2006Talia Porras SuarezNo ratings yet
- Metabolic Effects of Access To Sucrose DrinkDocument20 pagesMetabolic Effects of Access To Sucrose DrinkmineortizvNo ratings yet
- Current Basis and Future Directions of Zebrafish NutrigenomicsDocument10 pagesCurrent Basis and Future Directions of Zebrafish NutrigenomicsAlber AvendañoNo ratings yet
- Ovo e DCS MetabolicasDocument15 pagesOvo e DCS Metabolicasclissia croelhasNo ratings yet
- RESEARCH Open Access The Netherlands Cohort Study - Meat Investigation Cohort A Population-Based Cohort Over-Represented With Vegetarians, Pescetarians and Low Meat ConsumersDocument13 pagesRESEARCH Open Access The Netherlands Cohort Study - Meat Investigation Cohort A Population-Based Cohort Over-Represented With Vegetarians, Pescetarians and Low Meat ConsumersTonia RodriguesNo ratings yet
- Use of High-Fat Diets To Study Rodent Obesity As A Model of Human ObesityDocument2 pagesUse of High-Fat Diets To Study Rodent Obesity As A Model of Human Obesityinunk xiNo ratings yet
- Sex-Specific Differences in Hepatic Steatosis in Obese Spontaneously Hypertensive (SHROB) RatsDocument15 pagesSex-Specific Differences in Hepatic Steatosis in Obese Spontaneously Hypertensive (SHROB) RatsMarta SandeNo ratings yet
- Peanut Consumption and Cardiovascular RiskDocument6 pagesPeanut Consumption and Cardiovascular RiskWahyuningsih HamidNo ratings yet
- Pharmacology, Biochemistry and BehaviorDocument10 pagesPharmacology, Biochemistry and BehaviorYHOISS SMIHT MUNOZ CERONNo ratings yet
- Modeling Dietary Influences On Offspring Metabolic Programming in Drosophila MelanogasterDocument12 pagesModeling Dietary Influences On Offspring Metabolic Programming in Drosophila MelanogasterDaveMartoneNo ratings yet
- Pulse Crop Effects On Gut Microbial Populations, Intestinal Function, and Adiposity in A Mouse Model of Diet-Induced Obesity John N. McginleyDocument40 pagesPulse Crop Effects On Gut Microbial Populations, Intestinal Function, and Adiposity in A Mouse Model of Diet-Induced Obesity John N. Mcginleyzora.mendez637100% (8)
- Obesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationDocument10 pagesObesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationRodrigoNo ratings yet
- Dairy and Weight and Body CompositionDocument37 pagesDairy and Weight and Body CompositionSimone AdamiNo ratings yet
- Mediterranean-Style Diet in Pregnant Women With Metabolic Risk Factors (ESTEEM) : A Pragmatic Multicentre Randomised TrialDocument20 pagesMediterranean-Style Diet in Pregnant Women With Metabolic Risk Factors (ESTEEM) : A Pragmatic Multicentre Randomised Trialconstantius augustoNo ratings yet
- A Study of The Aphrodisiac Properties of Cordyceps Militaris in Streptozotocin-Induced Diabetic Male RatsDocument8 pagesA Study of The Aphrodisiac Properties of Cordyceps Militaris in Streptozotocin-Induced Diabetic Male RatsSami imasNo ratings yet
- Overfeeding Polyunsaturated and Saturated Fat Causes Distinct Effects On Liver and Visceral Fat Accumulation in HumansDocument13 pagesOverfeeding Polyunsaturated and Saturated Fat Causes Distinct Effects On Liver and Visceral Fat Accumulation in HumansnutrinathyalvesNo ratings yet
- Sremanakova Et Al-2018-Journal of Human Nutrition and DieteticsDocument11 pagesSremanakova Et Al-2018-Journal of Human Nutrition and DieteticsLulu BadriyahNo ratings yet
- Jensen2018 Article DietaryFatStimulatesDevelopmenDocument13 pagesJensen2018 Article DietaryFatStimulatesDevelopmenHijrawati Ayu WardaniNo ratings yet
- Effect of Long Term Consumption of Poultry Egg Products o - 2021 - Current DevelDocument13 pagesEffect of Long Term Consumption of Poultry Egg Products o - 2021 - Current DevelFelipe MoitaNo ratings yet
- Dairy Products and Health A Review of The Epidemiological EvidenceDocument6 pagesDairy Products and Health A Review of The Epidemiological EvidenceShell WattieNo ratings yet
- Bar-Maisels Et Al, 2017, Bone Lengh During Catch Up GrowthDocument16 pagesBar-Maisels Et Al, 2017, Bone Lengh During Catch Up GrowthCamila CajicaNo ratings yet
- Ayton 2020Document19 pagesAyton 2020Gaby ZavalaNo ratings yet
- Maternal High-Fat Diet Controls Offspring Kidney HDocument20 pagesMaternal High-Fat Diet Controls Offspring Kidney HManoj KumarNo ratings yet
- Wur Nutrition and Health ThesisDocument6 pagesWur Nutrition and Health Thesisgjgm36vk100% (2)
- Obesity - 2022 - Grech - Macronutrient Im Balance Drives Energy Intake in An Obesogenic Food Environment An EcologicalDocument12 pagesObesity - 2022 - Grech - Macronutrient Im Balance Drives Energy Intake in An Obesogenic Food Environment An EcologicalMelike KirişciNo ratings yet
- Hasil 1471-2458-14-1291Document6 pagesHasil 1471-2458-14-1291fathimzahroNo ratings yet
- Impact of Nutrition Counselling On Multiple Sclerosis Patients Nutritional Status: Randomized Controlled Clinical TrialDocument19 pagesImpact of Nutrition Counselling On Multiple Sclerosis Patients Nutritional Status: Randomized Controlled Clinical Trialmutiara fajriatiNo ratings yet
- Duodenal Endoluminal Barrier Sleeve Alters Gut Microbiota of ZDF RatsDocument28 pagesDuodenal Endoluminal Barrier Sleeve Alters Gut Microbiota of ZDF RatsIngrid MillanNo ratings yet
- Metabolic Diseases of Dairy Cattle - 2013 - VFP PDFDocument2 pagesMetabolic Diseases of Dairy Cattle - 2013 - VFP PDFfelipeNo ratings yet
- Summary: Sex-Dependent Effects of Cadmium Exposure in Early Life On Gut Microbiota and Fat Accumulation in MiceDocument3 pagesSummary: Sex-Dependent Effects of Cadmium Exposure in Early Life On Gut Microbiota and Fat Accumulation in Miceapi-296534570No ratings yet
- Translational Glycobiology in Human Health and DiseaseFrom EverandTranslational Glycobiology in Human Health and DiseaseMichelle KilcoyneNo ratings yet
- Diagnosis and Management of Hepatic Encephalopathy: A Case-based GuideFrom EverandDiagnosis and Management of Hepatic Encephalopathy: A Case-based GuideJasmohan S. BajajNo ratings yet
- Rius-Pérez 2020 PGC-1 Alfa, Inflammation, and Oxidative StressDocument20 pagesRius-Pérez 2020 PGC-1 Alfa, Inflammation, and Oxidative StressPaul SimononNo ratings yet
- Dupas 2017 MS and Hypertension Resulting From Fructose Enriched Diet in Wistar RatsDocument11 pagesDupas 2017 MS and Hypertension Resulting From Fructose Enriched Diet in Wistar RatsPaul SimononNo ratings yet
- Magge 2017 Edad en SMDocument14 pagesMagge 2017 Edad en SMPaul SimononNo ratings yet
- Gueugneau 2015 Skeletal Muscle Lipid Content and Oxidative Activity in Relation To Muscle Fiber Type in Aging and MSDocument12 pagesGueugneau 2015 Skeletal Muscle Lipid Content and Oxidative Activity in Relation To Muscle Fiber Type in Aging and MSPaul SimononNo ratings yet
- Cox 2011 Fructose Consumption and InflammationDocument5 pagesCox 2011 Fructose Consumption and InflammationPaul SimononNo ratings yet
- Rydén 2014 Adipose Tissue and Metabolic Alterations - Regional Differences in Fat Cell Size and Number Matter - But DifferentlyDocument7 pagesRydén 2014 Adipose Tissue and Metabolic Alterations - Regional Differences in Fat Cell Size and Number Matter - But DifferentlyPaul SimononNo ratings yet
- Uslusoy 2009 Liver Histology According To The Presence of Ms in NafldDocument6 pagesUslusoy 2009 Liver Histology According To The Presence of Ms in NafldPaul SimononNo ratings yet
- Vávrová 2013 Altered Activities of Antixodiant Enzymes in Patients With Metabolic SyndromeDocument9 pagesVávrová 2013 Altered Activities of Antixodiant Enzymes in Patients With Metabolic SyndromePaul SimononNo ratings yet
- Ohashi 2013 Association of MS With Kidney Fuction and Histology in Living Kidney DonorsDocument18 pagesOhashi 2013 Association of MS With Kidney Fuction and Histology in Living Kidney DonorsPaul SimononNo ratings yet
- Horii 2020 Islet Inflammation Is Associated With Pancreatic Fatty Infiltration and Hyperglycemia in Type 2 DiabetesDocument8 pagesHorii 2020 Islet Inflammation Is Associated With Pancreatic Fatty Infiltration and Hyperglycemia in Type 2 DiabetesPaul SimononNo ratings yet
- HORTELÃ - Ready To Drink Iced Teas From Microencapsulated Spearmint (Mentha Spicata L.) and Peppermint (Mentha Piperita L.) Extracts Physicochemical, Bioactive and Sensory CharacterizationDocument10 pagesHORTELÃ - Ready To Drink Iced Teas From Microencapsulated Spearmint (Mentha Spicata L.) and Peppermint (Mentha Piperita L.) Extracts Physicochemical, Bioactive and Sensory CharacterizationPaul SimononNo ratings yet
- Moreto 2014 The Higher Plasma MDA Concentrations Are Determined by MS Related GlucolipotoxicityDocument7 pagesMoreto 2014 The Higher Plasma MDA Concentrations Are Determined by MS Related GlucolipotoxicityPaul SimononNo ratings yet
- Laforest 2015 Adipocyte Size As A Determinant of Ms and Adipose Tissue DysfunctionDocument13 pagesLaforest 2015 Adipocyte Size As A Determinant of Ms and Adipose Tissue DysfunctionPaul SimononNo ratings yet
- Ramos-Romero 2016 Artículo OriginalDocument8 pagesRamos-Romero 2016 Artículo OriginalPaul SimononNo ratings yet
- Wang 2020 Novel Insights Into The Pathogenesis and Management of The MsDocument42 pagesWang 2020 Novel Insights Into The Pathogenesis and Management of The MsPaul SimononNo ratings yet
- Choudhary 2012 Correlation of Adipose Tissue With Liver Histology in Asian Indian Patients With Nonalcoholic Fatty Liver DiseaseDocument9 pagesChoudhary 2012 Correlation of Adipose Tissue With Liver Histology in Asian Indian Patients With Nonalcoholic Fatty Liver DiseasePaul SimononNo ratings yet
- Simopoulos 2013 Review Deficiencia de Omega 3Document23 pagesSimopoulos 2013 Review Deficiencia de Omega 3Paul SimononNo ratings yet
- Rodríguez-Monforte 2017 MS and Dietary PatternsDocument23 pagesRodríguez-Monforte 2017 MS and Dietary PatternsPaul SimononNo ratings yet
- Smith 2015 Epigenetic and Developmental Influences On The Risk of Obesity, Diabetes and MsDocument8 pagesSmith 2015 Epigenetic and Developmental Influences On The Risk of Obesity, Diabetes and MsPaul SimononNo ratings yet
- Jove 93 52172Document6 pagesJove 93 52172Paul SimononNo ratings yet
- Ferreira-Santos 2020 Artículo OriginalDocument10 pagesFerreira-Santos 2020 Artículo OriginalPaul SimononNo ratings yet
- Sil 2016 Artículo OriginalDocument10 pagesSil 2016 Artículo OriginalPaul SimononNo ratings yet
- He 2017 Gut Microbiota As A Potential Target of Ms - The Role of Probiotics and PrebioticsDocument14 pagesHe 2017 Gut Microbiota As A Potential Target of Ms - The Role of Probiotics and PrebioticsPaul SimononNo ratings yet
- ILGM SPECIES NI - Horsetails ScouringRushesDocument2 pagesILGM SPECIES NI - Horsetails ScouringRushesPaul SimononNo ratings yet
- Ketogenic Diet F3666Document1 pageKetogenic Diet F3666Paul SimononNo ratings yet
- Rodent Diet F1515 F0761Document1 pageRodent Diet F1515 F0761Paul SimononNo ratings yet
- Ye 2019 Lipotoxicity and Beta Cell Maintenance in Obesity and Type 2 DiabetesDocument15 pagesYe 2019 Lipotoxicity and Beta Cell Maintenance in Obesity and Type 2 DiabetesPaul SimononNo ratings yet
- Newsholme 2019 Oxidative Stress Pathways in Pancreatic Beta Cell and Insulin-Sensitive Cells and TissuesDocument14 pagesNewsholme 2019 Oxidative Stress Pathways in Pancreatic Beta Cell and Insulin-Sensitive Cells and TissuesPaul SimononNo ratings yet
- The Manufacture and Uses of Expanded Clay Aggregate: Thursday 15 November 2012 SCI HQ, LondonDocument36 pagesThe Manufacture and Uses of Expanded Clay Aggregate: Thursday 15 November 2012 SCI HQ, LondonVibhuti JainNo ratings yet
- ARK - Intel® Core™ I3-370m Processor (3M Cache, 2Document3 pagesARK - Intel® Core™ I3-370m Processor (3M Cache, 2Delzi Guindra AdriNo ratings yet
- Types of Loads Considered For Design of BridgeDocument45 pagesTypes of Loads Considered For Design of BridgeAbhishek100% (1)
- Book Index The Art of Heavy TransportDocument6 pagesBook Index The Art of Heavy TransportHermon Pakpahan50% (2)
- DJI F450 Construction Guide WebDocument21 pagesDJI F450 Construction Guide WebPutu IndrayanaNo ratings yet
- Reynold A. Nicholson - The Mystics of IslamDocument65 pagesReynold A. Nicholson - The Mystics of IslamLuminon SamanNo ratings yet
- Kami Export - Subject Complements-1 PDFDocument3 pagesKami Export - Subject Complements-1 PDFkcv kfdsaNo ratings yet
- Regression Analysis Random Motors ProjectDocument22 pagesRegression Analysis Random Motors ProjectPrateek AgrawalNo ratings yet
- Lima Indiana Oil FieldDocument32 pagesLima Indiana Oil FieldCHARLES PATULAYNo ratings yet
- 5 Contracting Activity and Technical Staff RequirementsDocument2 pages5 Contracting Activity and Technical Staff RequirementsDaniyar KussainovNo ratings yet
- Asaali - Project Estimation - Ce155p-2 - A73Document7 pagesAsaali - Project Estimation - Ce155p-2 - A73Kandhalvi AsaaliNo ratings yet
- Essential Calculus Skills Practice Workbook With Full SolutionsDocument528 pagesEssential Calculus Skills Practice Workbook With Full SolutionsGerardo Navarro Sánchez94% (65)
- The Working of KarmaDocument74 pagesThe Working of KarmaSuhas KulhalliNo ratings yet
- YoungMan EN131 GUIDEDocument16 pagesYoungMan EN131 GUIDErcpawar100% (1)
- Compiled LecsDocument24 pagesCompiled LecsNur SetsuNo ratings yet
- Warehouse Management Solution SheetDocument2 pagesWarehouse Management Solution Sheetpatelnandini109No ratings yet
- Kimi No Na Wa LibropdfDocument150 pagesKimi No Na Wa LibropdfSarangapani BorahNo ratings yet
- Chapter 7 - Stress - TransformationsDocument21 pagesChapter 7 - Stress - TransformationsroselleNo ratings yet
- Chap 4 - Basic Principles of ElectricityDocument31 pagesChap 4 - Basic Principles of ElectricityAdiseshuMiddeNo ratings yet
- Wcdma Idle Mode (Ericsson)Document29 pagesWcdma Idle Mode (Ericsson)Hosein ShahbaziNo ratings yet
- Haldex-Barnes 2-Stage Pump For Log SplittersDocument2 pagesHaldex-Barnes 2-Stage Pump For Log SplittersPer Akkamaan AgessonNo ratings yet
- Metaphysics of LucretiusDocument6 pagesMetaphysics of LucretiusChristopher BennettNo ratings yet
- Badminton ReviewerDocument10 pagesBadminton ReviewerHailsey WinterNo ratings yet
- Clinical Reviews: The Management of Children With Gastroenteritis and Dehydration in The Emergency DepartmentDocument13 pagesClinical Reviews: The Management of Children With Gastroenteritis and Dehydration in The Emergency DepartmentRina Dewi AnggraeniNo ratings yet
- PANASONIC - WebCam-TQZJ402 - TY-CC20W PDFDocument188 pagesPANASONIC - WebCam-TQZJ402 - TY-CC20W PDF123anthonyNo ratings yet
- Asco Series 238 ASCO Pilot Operated Solenoid Valves (Floating Diaphragm)Document2 pagesAsco Series 238 ASCO Pilot Operated Solenoid Valves (Floating Diaphragm)Khyle Laurenz DuroNo ratings yet
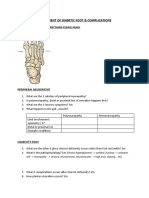
- Assessment of Diabetic FootDocument7 pagesAssessment of Diabetic FootChathiya Banu KrishenanNo ratings yet
- LinkageDocument9 pagesLinkageHarshu JunghareNo ratings yet
- Paper-Czechowski-Slow-strain-rate Stress Corrosion Testing of Welded Joints of Al-Mg AlloysDocument4 pagesPaper-Czechowski-Slow-strain-rate Stress Corrosion Testing of Welded Joints of Al-Mg Alloysjavo0128No ratings yet
- Conceptual Artist in Nigeria UNILAGDocument13 pagesConceptual Artist in Nigeria UNILAGAdelekan FortuneNo ratings yet