You might also like
- Custard, Pudddings, Mousses & SoufflesDocument6 pagesCustard, Pudddings, Mousses & SoufflesSubrata RouthNo ratings yet
- Cheng 2017 Ratas Jóvenes y Ratas Adultas High Fat DietDocument10 pagesCheng 2017 Ratas Jóvenes y Ratas Adultas High Fat DietPaul SimononNo ratings yet
- Salt Sucrose - Final011020Document21 pagesSalt Sucrose - Final011020vaishnavimadanNo ratings yet
- Sucralosa y RatonesDocument13 pagesSucralosa y RatonesafamhbkNo ratings yet
- 1949-498X 4 1limaxDocument89 pages1949-498X 4 1limaxMarta BioNo ratings yet
- Feeding Sodium Selenite and Nano-Selenium Stimulates Growth and Oxidation Resistance in BroilersDocument9 pagesFeeding Sodium Selenite and Nano-Selenium Stimulates Growth and Oxidation Resistance in BroilersCLAUDIO VICTORIANO FARFAN MERMANo ratings yet
- Hafez 2017Document46 pagesHafez 2017Thảo HàNo ratings yet
- Rabbirt SpirullinaDocument5 pagesRabbirt SpirullinaAyvz Martija LambioNo ratings yet
- Feijão e Cálculos RenaisDocument11 pagesFeijão e Cálculos RenaisEmily FerreiraNo ratings yet
- Hunter Gatherer Weight LossDocument8 pagesHunter Gatherer Weight LossrlrubenkingNo ratings yet
- Antihypertensive Effect of Quercetin in Rats Fed With A High Fat High Sucrose DietDocument8 pagesAntihypertensive Effect of Quercetin in Rats Fed With A High Fat High Sucrose DietGabriel Oliveira de SouzaNo ratings yet
- Yuyu Wang, Pao Xu, Zhijuan Nie, Quanjie Li, Nailin Shao, Jiancao Gao, and Gangchun XuDocument9 pagesYuyu Wang, Pao Xu, Zhijuan Nie, Quanjie Li, Nailin Shao, Jiancao Gao, and Gangchun Xudaniel cretuNo ratings yet
- Biochemical Changes in Lipid and Carbohydrate Metabolism in Obese Diabetic Male Albino RatsDocument8 pagesBiochemical Changes in Lipid and Carbohydrate Metabolism in Obese Diabetic Male Albino RatsHashim OmarNo ratings yet
- Coltherd Et Al 2021 BJN NOAEL P Sources Feline DietsDocument16 pagesColtherd Et Al 2021 BJN NOAEL P Sources Feline DietsannemabNo ratings yet
- tmp8C49 TMPDocument13 pagestmp8C49 TMPFrontiersNo ratings yet
- Animals 12 02959Document9 pagesAnimals 12 02959Andrea G.No ratings yet
- Long-Term Sucrosedrinking Causes Increased Body Weight and Glucose Intolerance in Normal Male Rats.Document6 pagesLong-Term Sucrosedrinking Causes Increased Body Weight and Glucose Intolerance in Normal Male Rats.mineortizvNo ratings yet
- Prevalence and Risk Factors of Kidney Stone-ArticleDocument6 pagesPrevalence and Risk Factors of Kidney Stone-ArticleReesky Nanda Rockn'rollNo ratings yet
- Chen, Et Al (2016)Document9 pagesChen, Et Al (2016)Kharisma N. PuspitasariNo ratings yet
- Zhang 2016Document28 pagesZhang 2016Julieth Viviana Rodriguez RincónNo ratings yet
- Effect of Sulfonamides and Thioureas On The Thyroid Gland and Basal MetabolismDocument25 pagesEffect of Sulfonamides and Thioureas On The Thyroid Gland and Basal MetabolismM SaqibNo ratings yet
- Pengaruh Asupan Berbagai Jenis Biji-Bijian Terhadap Kadar Asam Urat Pada Tikus Putih Jantan (Rattus Norvegicus)Document6 pagesPengaruh Asupan Berbagai Jenis Biji-Bijian Terhadap Kadar Asam Urat Pada Tikus Putih Jantan (Rattus Norvegicus)Lidya LatifatulNo ratings yet
- 1 s2.0 S002203021100213X MainDocument9 pages1 s2.0 S002203021100213X MainJuniClaudia13No ratings yet
- Antidiabetic Effect of Fermented (Millet) Supplement in Alloxan Induced Hyperglycemic Wistar RatsDocument4 pagesAntidiabetic Effect of Fermented (Millet) Supplement in Alloxan Induced Hyperglycemic Wistar Ratsmaster apotNo ratings yet
- 205225-Article Text-512137-1-10-20210329Document12 pages205225-Article Text-512137-1-10-20210329Martynas JNo ratings yet
- Pengaruh Asupan Berbagai Jenis Biji-Bijian Terhadap Kadar Asam Urat Pada Tikus Putih Jantan (Rattus Norvegicus)Document6 pagesPengaruh Asupan Berbagai Jenis Biji-Bijian Terhadap Kadar Asam Urat Pada Tikus Putih Jantan (Rattus Norvegicus)deviNo ratings yet
- Effects of Dietary Fibre Source On Microbiota Composition in The Large Intestine of Suckling PigletsDocument7 pagesEffects of Dietary Fibre Source On Microbiota Composition in The Large Intestine of Suckling Pigletsjohn pierreNo ratings yet
- Validation of A Novel Diagnostic Test For Assessing The Risk of Urinary Uric ACID CRYSTALLIZACIONDocument6 pagesValidation of A Novel Diagnostic Test For Assessing The Risk of Urinary Uric ACID CRYSTALLIZACIONMARIANNE CASTILLO ESCOBARNo ratings yet
- 10 1016@j Aquaculture 2016 09 005Document6 pages10 1016@j Aquaculture 2016 09 005Courtney FergusonNo ratings yet
- Nfs Journal: Fangfang Song, Jieun Oh, Kyungwon Lee, Mi Sook ChoDocument6 pagesNfs Journal: Fangfang Song, Jieun Oh, Kyungwon Lee, Mi Sook ChoClararida RiawanNo ratings yet
- Cytogenetic and Teratological Effects of MancozebDocument9 pagesCytogenetic and Teratological Effects of Mancozebforward websiteNo ratings yet
- Asian Paci Fic Journal of Tropical BiomedicineDocument5 pagesAsian Paci Fic Journal of Tropical BiomedicineAris Sandi PratamaNo ratings yet
- Effects of Crude Kerosene On Testosterone Levels, Aggression and Toxicity in RatDocument10 pagesEffects of Crude Kerosene On Testosterone Levels, Aggression and Toxicity in RatMevinNo ratings yet
- Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch PolysaccharidesDocument34 pagesShort-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharidesbdalcin5512No ratings yet
- Herbs For Gout: Alternative and Complementary Therapies October 2016Document9 pagesHerbs For Gout: Alternative and Complementary Therapies October 2016catalinNo ratings yet
- A Study of The Aphrodisiac Properties of Cordyceps Militaris in Streptozotocin-Induced Diabetic Male RatsDocument8 pagesA Study of The Aphrodisiac Properties of Cordyceps Militaris in Streptozotocin-Induced Diabetic Male RatsSami imasNo ratings yet
- Lindenschmidt 1991Document8 pagesLindenschmidt 1991aparna.cNo ratings yet
- The Effect of Aloin in Blood Glucose and Antioxidants in Male Albino Rats With Streptozotocin-Induced DiabeticDocument18 pagesThe Effect of Aloin in Blood Glucose and Antioxidants in Male Albino Rats With Streptozotocin-Induced DiabeticJunaid MinhasNo ratings yet
- 005 Sudirman2021 - Hippocampus Kuda Protein Hydrolysate Improves Male Reproductive - RedDocument9 pages005 Sudirman2021 - Hippocampus Kuda Protein Hydrolysate Improves Male Reproductive - RedSabri SudirmanNo ratings yet
- Effects of Saccharomyces Cerevisiae (Saccharomycetes: Saccharomycetaceae) On Astronotus Ocellatus As Growth Promoter and Immuno StimulantDocument13 pagesEffects of Saccharomyces Cerevisiae (Saccharomycetes: Saccharomycetaceae) On Astronotus Ocellatus As Growth Promoter and Immuno Stimulantmeliana rahayuNo ratings yet
- Efficacy of Sida Pilosa Aqueous Extract Against S Mansoni-Induced Granulomatous InflammationDocument15 pagesEfficacy of Sida Pilosa Aqueous Extract Against S Mansoni-Induced Granulomatous InflammationHermine Jatsa BoukengNo ratings yet
- Lactobacilli and Streptococcus Mutans in Saliva, Diet and Caries Increment in 8-And 13-Year-Old ChildrenDocument10 pagesLactobacilli and Streptococcus Mutans in Saliva, Diet and Caries Increment in 8-And 13-Year-Old ChildrenScorpioster KikiNo ratings yet
- Spirulina Reduces Diet-Induced Obesity Through Downregulation of Lipogenic Genes Expression inDocument10 pagesSpirulina Reduces Diet-Induced Obesity Through Downregulation of Lipogenic Genes Expression inMtra. Mayra N. Elizalde VillarrealNo ratings yet
- Effects of Topical Application of Lipopolysaccharide and Proteases On Hepatic Injury Induced by High-Cholesterol Diet in RatsDocument7 pagesEffects of Topical Application of Lipopolysaccharide and Proteases On Hepatic Injury Induced by High-Cholesterol Diet in RatsastrotactinNo ratings yet
- Summary: Sex-Dependent Effects of Cadmium Exposure in Early Life On Gut Microbiota and Fat Accumulation in MiceDocument3 pagesSummary: Sex-Dependent Effects of Cadmium Exposure in Early Life On Gut Microbiota and Fat Accumulation in Miceapi-296534570No ratings yet
- Measuring Parasitoid Carbohydrate Levels To Improve Biological ControlDocument5 pagesMeasuring Parasitoid Carbohydrate Levels To Improve Biological ControlBerton Swandi SipayungNo ratings yet
- Thaumatin Is Similar To Water in Blood Glucose Response in Wistar RatsDocument7 pagesThaumatin Is Similar To Water in Blood Glucose Response in Wistar RatsijasrjournalNo ratings yet
- Aquaculture: Yangen Zhou, Ramasamy Thirumurugan, Qingkui Wang, Chong M. Lee, D. Allen DavisDocument7 pagesAquaculture: Yangen Zhou, Ramasamy Thirumurugan, Qingkui Wang, Chong M. Lee, D. Allen DavisOdenkz XtreesidersNo ratings yet
- Steyn 2014Document3 pagesSteyn 2014RozavenaNo ratings yet
- Effect of Sodium Metabisulphite On Blood Metabolic Status of Wistar RatsDocument10 pagesEffect of Sodium Metabisulphite On Blood Metabolic Status of Wistar Ratsام محمدNo ratings yet
- DSFGDFG DDGHGHGGDocument402 pagesDSFGDFG DDGHGHGGRouf KashmiriNo ratings yet
- 1 s2.0 S1319562X23002358 MainDocument12 pages1 s2.0 S1319562X23002358 MainArif ElsanNo ratings yet
- Http://ocw Mit Edu/help/faq-TechnologyDocument17 pagesHttp://ocw Mit Edu/help/faq-TechnologySally ElshalqamyNo ratings yet
- 2020 Metabolic Risk FactorsDocument6 pages2020 Metabolic Risk FactorsAngela LópezNo ratings yet
- Najam TJPR Feb 2018Document6 pagesNajam TJPR Feb 2018Abdul SamadNo ratings yet
- Angelova 2013 Review Models of Obesity and Metabolic SyndromeDocument8 pagesAngelova 2013 Review Models of Obesity and Metabolic SyndromePaul SimononNo ratings yet
- Elsevier Scientific Publishers Ireland LTD.: Drug and Alcohol Dependence, 19 (1987) 333-344 333Document12 pagesElsevier Scientific Publishers Ireland LTD.: Drug and Alcohol Dependence, 19 (1987) 333-344 333Jef_8No ratings yet
- Acanthophora SpiciferaDocument8 pagesAcanthophora SpiciferaMuthya BrilianNo ratings yet
- Am J Physiol Regul Integr Comp Physiol-2001-Kristan-R502-10Document10 pagesAm J Physiol Regul Integr Comp Physiol-2001-Kristan-R502-10EstuardoANo ratings yet
- Manual of Animal AndrologyFrom EverandManual of Animal AndrologyPeter J ChenowethNo ratings yet
- Cyclospora and Cyclosporiasis: Epidemiology, Diagnosis, Detection, and ControlFrom EverandCyclospora and Cyclosporiasis: Epidemiology, Diagnosis, Detection, and ControlNo ratings yet
- Nutrition Mentalhealth Rao2008Document7 pagesNutrition Mentalhealth Rao2008Dinda Aulia HirdaNo ratings yet
- Lambert 2015Document9 pagesLambert 2015José Carlos Sánchez-RamirezNo ratings yet
- 8 fr-2018-220 Stephanie 1Document8 pages8 fr-2018-220 Stephanie 1nisceNo ratings yet
- Trace EvidentsDocument7 pagesTrace EvidentsnisceNo ratings yet
- Cyanidepaper 1Document12 pagesCyanidepaper 1nisceNo ratings yet
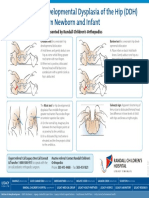
- 21 RandallChildrens CoMgmtGL Ortho HipDysplasia Illustrations CHC 4610 0217Document1 page21 RandallChildrens CoMgmtGL Ortho HipDysplasia Illustrations CHC 4610 0217nisceNo ratings yet
- Antidotes For Cyanide Poisoning.16Document2 pagesAntidotes For Cyanide Poisoning.16nisceNo ratings yet
- 1825-Article Text-6444-1-10-20210717Document11 pages1825-Article Text-6444-1-10-20210717nisceNo ratings yet
- Chain of Custody FormDocument1 pageChain of Custody FormnisceNo ratings yet
- 1 s2.0 S1877042812001048 MainDocument4 pages1 s2.0 S1877042812001048 MainnisceNo ratings yet
- (15435474 - Journal of Physical Activity and Health) Physical Inactivity and COVID-19 When Pandemics CollideDocument2 pages(15435474 - Journal of Physical Activity and Health) Physical Inactivity and COVID-19 When Pandemics CollidenisceNo ratings yet
- Tugas Buk Kasyani Jurnal Sistem Imun Dan ObesitasDocument5 pagesTugas Buk Kasyani Jurnal Sistem Imun Dan ObesitasfideNo ratings yet
- Sianida 1Document20 pagesSianida 1nisceNo ratings yet
- Sianida 2Document12 pagesSianida 2nisceNo ratings yet
- Consuming Highly Refined Carbohydrates Increases Risk of DepressionDocument2 pagesConsuming Highly Refined Carbohydrates Increases Risk of DepressionnisceNo ratings yet
- 816 820Document7 pages816 820PratiwiLaboroNo ratings yet
- Data BukuDocument28 pagesData BukunisceNo ratings yet
- Kadar Sianida Rokok Dan UdaraDocument2 pagesKadar Sianida Rokok Dan UdaranisceNo ratings yet
- What Are Carbohydrates?Document3 pagesWhat Are Carbohydrates?nisceNo ratings yet
- Free Illustration Resources: Use This Free Recolorable Icons and Illustrations in Your Canva DesignDocument18 pagesFree Illustration Resources: Use This Free Recolorable Icons and Illustrations in Your Canva DesignnisceNo ratings yet
- Calories ValueDocument103 pagesCalories ValueJ.n. AcharyaNo ratings yet
- Aerobic Vs Anaerobic Exercise Training Effects On The Cardiovascular SystemDocument6 pagesAerobic Vs Anaerobic Exercise Training Effects On The Cardiovascular SystemMohamad Gilang Harry HNo ratings yet
- Meal Timing Regulates The Human Circadian System: Highlights AuthorsDocument12 pagesMeal Timing Regulates The Human Circadian System: Highlights AuthorsnisceNo ratings yet
- Abnormal Food Timing and Circadian Dyssynchrony in Alcohol Induced Colon CarcinogenesisDocument20 pagesAbnormal Food Timing and Circadian Dyssynchrony in Alcohol Induced Colon CarcinogenesisnisceNo ratings yet
- Measles During Arbovirus Outbreak: A Diagnostic Challenge: Case ReportDocument4 pagesMeasles During Arbovirus Outbreak: A Diagnostic Challenge: Case ReportnisceNo ratings yet
- Hubungan Masa Kerja, Lama Kerja, Lama Penyemprotan Dan Frekuensi Penyemprotan Terhadap Kadar Kolinesterase Dalam Darah Pada Petani Di Desa Sumberejo Kecamatan Ngablak Kabupaten MagelangDocument7 pagesHubungan Masa Kerja, Lama Kerja, Lama Penyemprotan Dan Frekuensi Penyemprotan Terhadap Kadar Kolinesterase Dalam Darah Pada Petani Di Desa Sumberejo Kecamatan Ngablak Kabupaten MagelangnisceNo ratings yet
- Class Indications: GriseofulvinDocument1 pageClass Indications: GriseofulvinnisceNo ratings yet
- About My Self About My Self: From Independent University of BangladeshDocument2 pagesAbout My Self About My Self: From Independent University of BangladeshMd. Hafijur Rahman PlabonNo ratings yet
- Management PrinciplesDocument12 pagesManagement PrinciplesminminNo ratings yet
- 4th-Quarter - WEEK - 2 - Counting The Number of Occurrences of An Outcome in An ExperimentDocument24 pages4th-Quarter - WEEK - 2 - Counting The Number of Occurrences of An Outcome in An ExperimentErika Lloren Luyun-GaliaNo ratings yet
- Rice 1999 Pottery OriginsDocument54 pagesRice 1999 Pottery OriginsyaneliNo ratings yet
- Animal CareDocument12 pagesAnimal Caremirzaayan918No ratings yet
- Essays On PovertyDocument7 pagesEssays On Povertydnrlbgnbf100% (1)
- Artefacts in Conventional Scanning Electron Microscopy of Some MiDocument18 pagesArtefacts in Conventional Scanning Electron Microscopy of Some Mijohnbender35814163No ratings yet
- T6 Mrs. M. L. BhavyaDocument4 pagesT6 Mrs. M. L. BhavyaHet BhaljaNo ratings yet
- Congres USAMV Book of Abstracts 2018 MilleniumDocument413 pagesCongres USAMV Book of Abstracts 2018 MilleniumVlad TomaNo ratings yet
- Essential Oils As Antioxidants: Their Evaluation by DPPH, ABTS, Frap, Cuprac, and B-Carotene Bleaching MethodsDocument9 pagesEssential Oils As Antioxidants: Their Evaluation by DPPH, ABTS, Frap, Cuprac, and B-Carotene Bleaching MethodsVincentius KevinNo ratings yet
- Experiment No. 3 Heat of Reaction: 4.18 X 10 KJ/G 4.18 X 10 KJ/GDocument3 pagesExperiment No. 3 Heat of Reaction: 4.18 X 10 KJ/G 4.18 X 10 KJ/GjohannNo ratings yet
- ES MANUAL Cafetera Bomba Tradicional Espresso EC270 DelonghiDocument17 pagesES MANUAL Cafetera Bomba Tradicional Espresso EC270 Delonghialejandro ferrer0% (1)
- Capstone Group6 Stem6Document23 pagesCapstone Group6 Stem6Yuan Andrei SantosNo ratings yet
- Exporters Directory Province:Jawa Timur Product:Frozen Raw ShrimpDocument2 pagesExporters Directory Province:Jawa Timur Product:Frozen Raw ShrimpV.A. Elvien Kristian PrasetyoNo ratings yet
- Ssac 2104Document100 pagesSsac 2104Amrutha amrNo ratings yet
- Cookery-9-Q4m3 - Glaiza R LadimoDocument16 pagesCookery-9-Q4m3 - Glaiza R LadimoMichelle Del Rosario MercadoNo ratings yet
- Cotton FibreDocument26 pagesCotton Fibrekavi RAJ SenNo ratings yet
- Earth AND Life Science: Worksheet/S On Genetic Engineering and Animal Organ SystemDocument8 pagesEarth AND Life Science: Worksheet/S On Genetic Engineering and Animal Organ SystemBei CabNo ratings yet
- Food Premises Design Construction and Fit Out GuideDocument55 pagesFood Premises Design Construction and Fit Out GuideЕлена КоваленкоNo ratings yet
- 4000 Essential English Words 3Document197 pages4000 Essential English Words 3apaxfranklin.comNo ratings yet
- Tugas Ing Wajib No Absen 1Document5 pagesTugas Ing Wajib No Absen 1Yunisha MaulidyanaNo ratings yet
- SOAL Latihan PAS B.Inggris, Grade 8Document7 pagesSOAL Latihan PAS B.Inggris, Grade 8blushNo ratings yet
- Lesson E: Have You Been ToDocument2 pagesLesson E: Have You Been ToKARLA PAOLA CURIEL MONTUFARNo ratings yet
- Prac - Research EeeeeeeeDocument16 pagesPrac - Research EeeeeeeeMarry Ann luboNo ratings yet
- Pat Big KLS 9 Genap 22-23Document4 pagesPat Big KLS 9 Genap 22-23khuluk payamanNo ratings yet
- Whitelabel Rate ListDocument1 pageWhitelabel Rate ListBiotech FormularyNo ratings yet
- Chemical Composition and Antioxidant Activities of Black Seed Oil (Nigella Sativa L.)Document7 pagesChemical Composition and Antioxidant Activities of Black Seed Oil (Nigella Sativa L.)Dian Takwa HarahapNo ratings yet
- Result 3 For 'Word Power'Document69 pagesResult 3 For 'Word Power'elsa rahayuNo ratings yet
- BOTH, NEITHER and EITHERDocument2 pagesBOTH, NEITHER and EITHERIrma GochaleishviliNo ratings yet