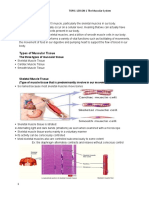
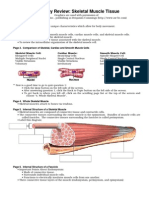
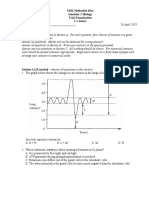
You might also like
- Excitation Contraction CouplingDocument34 pagesExcitation Contraction CouplingRUdraNo ratings yet
- Chapter 12 - Muscle PhysiologyDocument21 pagesChapter 12 - Muscle PhysiologyAlec McIntoshNo ratings yet
- MUSCLESDocument3 pagesMUSCLESalihusseinNo ratings yet
- Integrated SGDDocument10 pagesIntegrated SGDALDOUS VINCENT MENDOZANo ratings yet
- Module 10 Kin 267 Muscle TissueDocument4 pagesModule 10 Kin 267 Muscle TissueJ aNo ratings yet
- Skeletal Muscle Physiology LectureDocument65 pagesSkeletal Muscle Physiology Lectureanon_737973908No ratings yet
- Biochemistry of MuscleDocument68 pagesBiochemistry of MuscleWisnu KuncoroNo ratings yet
- Smooth Muscle Physiology by Bushra Akhtar Dhillon (07.04.2020)Document24 pagesSmooth Muscle Physiology by Bushra Akhtar Dhillon (07.04.2020)Laraib SohailNo ratings yet
- G. Muscle Physiology - 1Document45 pagesG. Muscle Physiology - 1Milimo MweembaNo ratings yet
- Muscoskeletal SystemDocument5 pagesMuscoskeletal SystemJeevikaGoyalNo ratings yet
- Exercise PhysiologyDocument2 pagesExercise PhysiologyHya LaniogNo ratings yet
- Muscles 1Document25 pagesMuscles 1Mihaela CosteaNo ratings yet
- Muscles1Document25 pagesMuscles1sairash.khan23No ratings yet
- Coordination: Physiology of Muscle Contraction and Different Muscle Contraction (Isotonic, Isometric, Isokinetic)Document6 pagesCoordination: Physiology of Muscle Contraction and Different Muscle Contraction (Isotonic, Isometric, Isokinetic)nn1216No ratings yet
- General-Physiology - Smooth MuscleDocument4 pagesGeneral-Physiology - Smooth Muscledagehi6880No ratings yet
- By Dr.H.Gusbakti, MD, MSC, PKK, AIFM Professor of Physiology University Islamic of North SumateraDocument83 pagesBy Dr.H.Gusbakti, MD, MSC, PKK, AIFM Professor of Physiology University Islamic of North SumateraJennifer WootenNo ratings yet
- The Muscular System: B. Pimentel, M.D. University of Makati - College of NursingDocument71 pagesThe Muscular System: B. Pimentel, M.D. University of Makati - College of Nursingapi-19824701No ratings yet
- Physiology of Excitable Tissue and Action PotentialsDocument12 pagesPhysiology of Excitable Tissue and Action PotentialsTamar PertiaNo ratings yet
- SGD 4 - Skeletal Muscle PhysiologyDocument3 pagesSGD 4 - Skeletal Muscle PhysiologyKriska Noelle0% (1)
- Kuliah Muscle Contraction 2010Document69 pagesKuliah Muscle Contraction 2010bagir_dm10No ratings yet
- Smooth Muscle Contraction and Relaxation Advances in Physiology Education PDFDocument1 pageSmooth Muscle Contraction and Relaxation Advances in Physiology Education PDFBrizo BNo ratings yet
- MSS Module Year One 2024Document43 pagesMSS Module Year One 2024Thaw Tar Noe WaiNo ratings yet
- Skeletal Muscle Contraction MechanismDocument4 pagesSkeletal Muscle Contraction MechanismAnn SoonNo ratings yet
- Lecture 4Document6 pagesLecture 4Mahreen NoorNo ratings yet
- 09smooth MuscleDocument13 pages09smooth MuscleJaydave PatelNo ratings yet
- Short Essay QuestionsDocument2 pagesShort Essay QuestionsMustajab AlizaNo ratings yet
- Mean While,: ("Center" or "Circle") - So, Concentric Things Have A Center in Common. (Document4 pagesMean While,: ("Center" or "Circle") - So, Concentric Things Have A Center in Common. (Karylle SequinaNo ratings yet
- Skeletal Muscle Contraction via Sliding Filament MechanismDocument3 pagesSkeletal Muscle Contraction via Sliding Filament MechanismAJ AbraganNo ratings yet
- Excitation PrintDocument3 pagesExcitation PrintZoRo4everNo ratings yet
- L-3 - Locomotion and Movement - Sept 30, 2019 PDFDocument45 pagesL-3 - Locomotion and Movement - Sept 30, 2019 PDFKent Wells100% (1)
- Initiation of Muscle Contraction MuscleDocument3 pagesInitiation of Muscle Contraction MuscleManu JamesNo ratings yet
- 38.18 Muscle Contraction and Locomotion - Regulatory ProteinsDocument2 pages38.18 Muscle Contraction and Locomotion - Regulatory ProteinsRejoice OtungNo ratings yet
- Muscles 1Document25 pagesMuscles 1Catalina ZamfirNo ratings yet
- Muscle Physiology Lecture on Neuromuscular Junction and Contraction MechanismsDocument20 pagesMuscle Physiology Lecture on Neuromuscular Junction and Contraction MechanismsSandy MonirNo ratings yet
- The Sliding Filament Mechanism of Muscle ContractionDocument2 pagesThe Sliding Filament Mechanism of Muscle ContractionShelene KarriedoNo ratings yet
- September 2, 2016 Muscle 1Document37 pagesSeptember 2, 2016 Muscle 1bluemoon4777No ratings yet
- ASSIGNMENT 2 Physiology 2Document3 pagesASSIGNMENT 2 Physiology 2jana7-7No ratings yet
- (Review) Winter Term Test Review AnswersDocument5 pages(Review) Winter Term Test Review AnswersJamie SamuelNo ratings yet
- Sliding Filament Theory of Muscle ContractionDocument5 pagesSliding Filament Theory of Muscle ContractionRica NorcioNo ratings yet
- Physiology Linda Costanzo 6th 1Document50 pagesPhysiology Linda Costanzo 6th 1Moonnime BlueNo ratings yet
- Muscle Physiology: A Guide to Muscle Structure and Contraction MechanismsDocument20 pagesMuscle Physiology: A Guide to Muscle Structure and Contraction MechanismsPisiform90100% (1)
- Biophysical Events of Contraction-Relaxation in Muscle Tissue - AcikdersDocument8 pagesBiophysical Events of Contraction-Relaxation in Muscle Tissue - AcikdersErde NmuNo ratings yet
- Cross Bridge CycleDocument14 pagesCross Bridge CycleIdenyi Daniel Ewa EdeNo ratings yet
- Chapter 12 Muscle Physiology - Important Questions: 6. Mechanism of Muscle ContractionDocument7 pagesChapter 12 Muscle Physiology - Important Questions: 6. Mechanism of Muscle ContractionIrum RafeeqNo ratings yet
- Smooth MuscleDocument8 pagesSmooth Muscleصفا جواد كاظمNo ratings yet
- Bche 4090 1155168548Document7 pagesBche 4090 1155168548Yenny TsaiNo ratings yet
- Mechanism of Muscle ContractionDocument24 pagesMechanism of Muscle Contractionfisika100% (1)
- Instuderings Q5Document2 pagesInstuderings Q5Nora SöderbergNo ratings yet
- Myosin Powered Movement and Contraction in Muscle and Non-Muscle CellsDocument8 pagesMyosin Powered Movement and Contraction in Muscle and Non-Muscle CellsIndranil BanerjeeNo ratings yet
- Sliding Filament Theory Sem 5 AssignmentDocument2 pagesSliding Filament Theory Sem 5 Assignmentparth choudharyNo ratings yet
- 15 Control and CoordinationDocument6 pages15 Control and CoordinationKevin XingNo ratings yet
- Control of Skeletal Muscle Contractions ExplainedDocument3 pagesControl of Skeletal Muscle Contractions ExplainedTofik MohammedNo ratings yet
- Muscle Contraction MechanismDocument14 pagesMuscle Contraction MechanismAlexNo ratings yet
- MuscleDocument8 pagesMuscleJonabel CubolNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Bio 336 Physio Exam 1 EssaysDocument3 pagesBio 336 Physio Exam 1 EssayssheilaNo ratings yet
- Physiology of Muscle Contraction and Control of MovementDocument69 pagesPhysiology of Muscle Contraction and Control of MovementMakmunNawil100% (1)
- Skeletal Muscle Physiology ExplainedDocument26 pagesSkeletal Muscle Physiology ExplainedUSAMA AHMEDNo ratings yet
- DhruvalHow Is Strength of Contraction Regulated in Different Types of MuscleDocument5 pagesDhruvalHow Is Strength of Contraction Regulated in Different Types of MuscleDhruvalNo ratings yet
- Q 1Document33 pagesQ 1m1.nourianNo ratings yet
- Lore of Running 4th Ed PDFDocument946 pagesLore of Running 4th Ed PDFwooyann64% (11)
- Bank F MCQS: Etea Medical PAPER 2019Document14 pagesBank F MCQS: Etea Medical PAPER 2019asifNo ratings yet
- Bio201 Exam 2 Master With AnswersDocument12 pagesBio201 Exam 2 Master With AnswersRicha Rala100% (1)
- Ch11 Trans Membrane Transport of Ions and Small MoleculeDocument113 pagesCh11 Trans Membrane Transport of Ions and Small MoleculeamuthukumaranNo ratings yet
- PHYSIO Cardiac Muscle and Heart As A PumpDocument5 pagesPHYSIO Cardiac Muscle and Heart As A PumpAyumi StarNo ratings yet
- A Practical Approach To Cardiac Anesthesia-Lippincot - Wolters Kluwer (2012)Document1,677 pagesA Practical Approach To Cardiac Anesthesia-Lippincot - Wolters Kluwer (2012)ElenaCondratscribdNo ratings yet
- Group VII - Smooth Muscle ContractionDocument7 pagesGroup VII - Smooth Muscle ContractioniqbalNo ratings yet
- Official Bank-MusclesDocument27 pagesOfficial Bank-MusclesEbtihal AlharthiNo ratings yet
- Anaphy Muscular SystemDocument54 pagesAnaphy Muscular SystemZosia SageNo ratings yet
- Physiology. Hrd.Document549 pagesPhysiology. Hrd.Sʌɩĸʌt PʌʋɭNo ratings yet
- 397 - Musculoskeletal Physiology) Smooth MuscleDocument5 pages397 - Musculoskeletal Physiology) Smooth Musclevkpremiyahoo.comNo ratings yet
- Drugs Used in Heart Failure: Pharmacology (2) PHAR 342Document19 pagesDrugs Used in Heart Failure: Pharmacology (2) PHAR 342Dana HamarshehNo ratings yet
- 5 Measurement of Water-Holding Capacity and Juiciness: K.O. Honikel and R. HammDocument37 pages5 Measurement of Water-Holding Capacity and Juiciness: K.O. Honikel and R. HammFrancisco QuiñonesNo ratings yet
- Skeletal Muscle Anatomy ReviewDocument6 pagesSkeletal Muscle Anatomy ReviewKwai Fong ChanNo ratings yet
- P2 2019 SMK Methodist PDFDocument11 pagesP2 2019 SMK Methodist PDFBryanLeeChienYungNo ratings yet
- Berg 8e Testbank Chapter13 PDFDocument13 pagesBerg 8e Testbank Chapter13 PDFbenin2No ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- Understanding Muscle Contraction at the Molecular LevelDocument30 pagesUnderstanding Muscle Contraction at the Molecular LevelMaggie-Louise BellamyNo ratings yet
- Muscle Tissue NotesDocument11 pagesMuscle Tissue NotesAubrey Unique EvangelistaNo ratings yet
- Muscle Physio PDFDocument41 pagesMuscle Physio PDFCamille MagdirilaNo ratings yet
- Chapter Six The Muscular SystemDocument30 pagesChapter Six The Muscular SystemabiyotNo ratings yet
- Questionpaper Unit5 (WBI05) June2017Document28 pagesQuestionpaper Unit5 (WBI05) June2017Rabia HasanNo ratings yet
- 3 Gen Beta BlockersDocument10 pages3 Gen Beta Blockersgnaneshwar nNo ratings yet
- Sim Muscle TutorialDocument20 pagesSim Muscle TutorialKiran KumarNo ratings yet
- The Enzyme List Class 7 - Translocases: ExplorenzDocument35 pagesThe Enzyme List Class 7 - Translocases: ExplorenzgustasconNo ratings yet
- Thesis Michiel Maarten SteinzDocument79 pagesThesis Michiel Maarten SteinzShukr Wesman BlbasNo ratings yet
- Rhabdomyolysis ReviewDocument16 pagesRhabdomyolysis Reviewdavid_banner1No ratings yet
- G. Muscle Physiology - 1Document45 pagesG. Muscle Physiology - 1Milimo MweembaNo ratings yet
- Guyton Hall PHYSIOLOGY Chapter 8 PDFDocument7 pagesGuyton Hall PHYSIOLOGY Chapter 8 PDFOsman NazirNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)