You might also like
- Downsi 6 PDFDocument11 pagesDownsi 6 PDFmutimutimutiNo ratings yet
- Edrv 0439Document13 pagesEdrv 0439fasfaNo ratings yet
- Adiponectin in Diabetes Mellitus: N. Xita and A. TsatsoulisDocument8 pagesAdiponectin in Diabetes Mellitus: N. Xita and A. TsatsoulisDzil FikriNo ratings yet
- Article-2012-Vaspin in Obesity and Diabetes-Pathophysiological and Clinical SignificanceDocument7 pagesArticle-2012-Vaspin in Obesity and Diabetes-Pathophysiological and Clinical Significancecarlos ArozamenaNo ratings yet
- 29126.1 20150227101459 CoveredDocument10 pages29126.1 20150227101459 CoveredeeeeNo ratings yet
- Adipokinas AntiinflamatoriasDocument8 pagesAdipokinas AntiinflamatoriasArturo PerezNo ratings yet
- J. Biol. Chem.-2014-Zhao-29112-22Document12 pagesJ. Biol. Chem.-2014-Zhao-29112-22fatimakh462No ratings yet
- Fruta de TemporadaDocument10 pagesFruta de TemporadaERMESNo ratings yet
- 1340 FullDocument9 pages1340 FullAkma RahaNo ratings yet
- 2010 Article 9216 PDFDocument8 pages2010 Article 9216 PDFphoechoexNo ratings yet
- 10 1038@ncpgasthep0186Document8 pages10 1038@ncpgasthep0186Ronald Echeverria IbazetaNo ratings yet
- GLP-1 Elicits An Intrinsic Gut-Liver Metabolic Signal To Ameliorate Diet-Induced VLDL Overproduction and Insulin ResistanceDocument8 pagesGLP-1 Elicits An Intrinsic Gut-Liver Metabolic Signal To Ameliorate Diet-Induced VLDL Overproduction and Insulin ResistanceAnditri WeningtyasNo ratings yet
- Peptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. BraydenDocument7 pagesPeptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. Braydensuci apsari pebriantiNo ratings yet
- LOW ADIPONECTIN AND TYPE 2 DIABETESDocument5 pagesLOW ADIPONECTIN AND TYPE 2 DIABETESRahadiyan HadinataNo ratings yet
- Jurnal Covid Dan ObesitasDocument15 pagesJurnal Covid Dan ObesitasIcha DestiaNo ratings yet
- Moderate Weight Loss Attenuates Chronic Endoplasmic Reticulum Stress and Mitochondrial Dysfunction in Human ObesityDocument10 pagesModerate Weight Loss Attenuates Chronic Endoplasmic Reticulum Stress and Mitochondrial Dysfunction in Human ObesityJulia SCNo ratings yet
- The Integrative Biology of Type 2 DiabetesDocument10 pagesThe Integrative Biology of Type 2 DiabetesAdrian GhiţăNo ratings yet
- Adipose Tissue NAD and Biology in Obesity and Insulin Resistance - From Mechanism To TherapyDocument19 pagesAdipose Tissue NAD and Biology in Obesity and Insulin Resistance - From Mechanism To TherapyThamyres CristhinaNo ratings yet
- Evolutionary Benefit of Insulin ResistanceDocument6 pagesEvolutionary Benefit of Insulin ResistanceFrancielle A.RNo ratings yet
- Ando Adiponectin PresentationDocument22 pagesAndo Adiponectin PresentationNatashaa DiyaNo ratings yet
- Scheja & Heeren 2019 The Endocrine Function of Adipose Tissues in Health and Cardiometabolic DiseaseDocument18 pagesScheja & Heeren 2019 The Endocrine Function of Adipose Tissues in Health and Cardiometabolic DiseasevibhutiNo ratings yet
- La ResistinaDocument9 pagesLa Resistina4ndr3s.l EmaNo ratings yet
- Artículo JC Tema 9Document9 pagesArtículo JC Tema 9Margarita CadenaNo ratings yet
- Low Adiponectin Levels in Adolescent Obesity: A Marker of Increased Intramyocellular Lipid AccumulationDocument5 pagesLow Adiponectin Levels in Adolescent Obesity: A Marker of Increased Intramyocellular Lipid Accumulationsario indriayaniNo ratings yet
- NIH Public Access: Author ManuscriptDocument10 pagesNIH Public Access: Author ManuscriptErdelyi-Molnár ImolaNo ratings yet
- Central Chronic Apelin Infusion Decreases Energy Expenditure and Thermogenesis in MiceDocument12 pagesCentral Chronic Apelin Infusion Decreases Energy Expenditure and Thermogenesis in Micecarlos marcelo concha vilcaNo ratings yet
- Obesity & Weight Loss Therapy: Role of Leptin in ObesityDocument3 pagesObesity & Weight Loss Therapy: Role of Leptin in ObesityMiltonLondoñoNo ratings yet
- PA Induced InsulinDocument7 pagesPA Induced Insulinsajad ahmad MalikNo ratings yet
- Birkenfield 2014Document11 pagesBirkenfield 2014Bruno MoraesNo ratings yet
- Zhou 2019Document11 pagesZhou 2019Andreea Claudia NegrilaNo ratings yet
- Nej MR A 2216691Document15 pagesNej MR A 2216691lakshminivas PingaliNo ratings yet
- PEDF Regulates Lipid Metabolism Via ATGLDocument9 pagesPEDF Regulates Lipid Metabolism Via ATGLAngelNo ratings yet
- Defectos MetabólicosDocument14 pagesDefectos Metabólicosandres herreraNo ratings yet
- Leptin Resistance and Obesity: Pablo J. Enriori, Anne E. Evans, Puspha Sinnayah, and Michael A. CowleyDocument5 pagesLeptin Resistance and Obesity: Pablo J. Enriori, Anne E. Evans, Puspha Sinnayah, and Michael A. CowleyAlfa RidziNo ratings yet
- Reduced hepatic IDE activity contributes to hyperinsulinemia in glucocorticoid-treated rodentsDocument8 pagesReduced hepatic IDE activity contributes to hyperinsulinemia in glucocorticoid-treated rodentsFuada Haerana RifaiNo ratings yet
- Rohrbach, S 2007 AdiponeDocument35 pagesRohrbach, S 2007 AdiponeYHOISS SMIHT MUNOZ CERONNo ratings yet
- Stilbenes May Reduce Obesity by Affecting Adipogenesis, Lipogenesis, Lipolysis & ThermogenesisDocument8 pagesStilbenes May Reduce Obesity by Affecting Adipogenesis, Lipogenesis, Lipolysis & ThermogenesisTony Miguel Saba SabaNo ratings yet
- Basic and Translational - LiverDocument26 pagesBasic and Translational - Liverdr. Nur'aini HasanNo ratings yet
- Tjokroprawiro, 2005Document7 pagesTjokroprawiro, 2005Nadya SaptarinaNo ratings yet
- cphy.c190031Document37 pagescphy.c190031Javier AbalNo ratings yet
- Matsunami Et Al., 2010 My 5th PaparDocument10 pagesMatsunami Et Al., 2010 My 5th PaparTokio MatsunamiNo ratings yet
- Salidroside and Curcumin Formula Prevents Liver Injury in Nonalcoholic Fatty Liver Disease in RatsDocument10 pagesSalidroside and Curcumin Formula Prevents Liver Injury in Nonalcoholic Fatty Liver Disease in Ratsbagus aldiNo ratings yet
- High-Fat Diet Induces Endothelial Dysfunction Through Downregulation of the AMPK-PI3K-Akt-eNOS PathwayDocument13 pagesHigh-Fat Diet Induces Endothelial Dysfunction Through Downregulation of the AMPK-PI3K-Akt-eNOS PathwayJorge CamargoNo ratings yet
- Mechanisms of Insulin Action and Insulin Resistance (Petersen y Shulman, 2018)Document91 pagesMechanisms of Insulin Action and Insulin Resistance (Petersen y Shulman, 2018)NICOLÁS ANDRÉS AYELEF PARRAGUEZNo ratings yet
- Aarr 2008 10Document16 pagesAarr 2008 10lucccasssNo ratings yet
- Afadin phosphorylation negatively regulates insulin signaling in adipose tissueDocument16 pagesAfadin phosphorylation negatively regulates insulin signaling in adipose tissueEmil AndersenNo ratings yet
- Grupo 3 Obesity InflammationDocument9 pagesGrupo 3 Obesity InflammationJeisonAlejandroÁlvarezVergaraNo ratings yet
- Oliveiraandrade 2014Document8 pagesOliveiraandrade 2014JuniorMartinsNo ratings yet
- Biloia de Los PeroxisomasDocument8 pagesBiloia de Los PeroxisomasEdgar GalvánNo ratings yet
- Article: Correspondence: Lrobinso@uoguelph - Ca Tel.: +1-519-824-4120 (Ext. 52297)Document16 pagesArticle: Correspondence: Lrobinso@uoguelph - Ca Tel.: +1-519-824-4120 (Ext. 52297)Alma AcevedoNo ratings yet
- Original Article Apelin Inhibits Diet-Induced Obesity by Enhancing Lymphatic and Blood Vessel IntegrityDocument46 pagesOriginal Article Apelin Inhibits Diet-Induced Obesity by Enhancing Lymphatic and Blood Vessel IntegrityAnupam BhardwajNo ratings yet
- A Review On New Target Insulysine (Ide) & Its Inhibitors As Antidiabetics AgentsDocument12 pagesA Review On New Target Insulysine (Ide) & Its Inhibitors As Antidiabetics AgentsTJPRC PublicationsNo ratings yet
- Effects of Intermittent Fasting On Health, Aging, and DiseaseDocument11 pagesEffects of Intermittent Fasting On Health, Aging, and DiseaseGonzalo TabaresNo ratings yet
- Physiology, Endocrine, Pancreas - StatPearls - NCBI BookshelfDocument5 pagesPhysiology, Endocrine, Pancreas - StatPearls - NCBI Bookshelfdrpaez27No ratings yet
- Fang 2005Document10 pagesFang 2005pamela gaitanNo ratings yet
- Possible Pharmacological Approach Targeting Endoplasmic Reticulum Stress To Ameliorate Leptin Resistance in ObesityDocument4 pagesPossible Pharmacological Approach Targeting Endoplasmic Reticulum Stress To Ameliorate Leptin Resistance in ObesitySyahrul TubaNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument8 pagesBiochemical and Biophysical Research CommunicationsAnnisa khairaNo ratings yet
- AMPc y MetaboismoDocument8 pagesAMPc y MetaboismoEmilio MoncadaNo ratings yet
- Ayuno IntermitenteDocument11 pagesAyuno IntermitenteNancyRojo3gmail.comNo ratings yet
- Apelin and Energy Metabolism: Chantal Bertrand, Philippe Valet and Isabelle Castan-LaurellDocument5 pagesApelin and Energy Metabolism: Chantal Bertrand, Philippe Valet and Isabelle Castan-LaurellErdelyi-Molnár ImolaNo ratings yet
- 10.1016/j.vph.2015.08.005: Vascular PharmacologyDocument24 pages10.1016/j.vph.2015.08.005: Vascular PharmacologyErdelyi-Molnár ImolaNo ratings yet
- European Journal of Pharmacology: C. Chaves-Almagro, I. Castan-Laurell, C. Dray, C. Knauf, P. Valet, B. MasriDocument11 pagesEuropean Journal of Pharmacology: C. Chaves-Almagro, I. Castan-Laurell, C. Dray, C. Knauf, P. Valet, B. MasriErdelyi-Molnár ImolaNo ratings yet
- Apelin, Elabela/Toddler, and Biased Agonists As Novel Therapeutic Agents in The Cardiovascular SystemDocument8 pagesApelin, Elabela/Toddler, and Biased Agonists As Novel Therapeutic Agents in The Cardiovascular SystemErdelyi-Molnár ImolaNo ratings yet
- The Apelin Receptor: Physiology, Pathology, Cell Signalling, and Ligand Modulation of A Peptide-Activated Class A GPCRDocument21 pagesThe Apelin Receptor: Physiology, Pathology, Cell Signalling, and Ligand Modulation of A Peptide-Activated Class A GPCRErdelyi-Molnár ImolaNo ratings yet
- Clinica Chimica ActaDocument8 pagesClinica Chimica ActaErdelyi-Molnár ImolaNo ratings yet
- Apelin Attenuates Oxidative Stress in Human AdipocytesDocument12 pagesApelin Attenuates Oxidative Stress in Human AdipocytesErdelyi-Molnár ImolaNo ratings yet
- Apelin and ACE2 in Cardiovascular Disease: Anastasia Z Kalea & Daniel BatlleDocument10 pagesApelin and ACE2 in Cardiovascular Disease: Anastasia Z Kalea & Daniel BatlleErdelyi-Molnár ImolaNo ratings yet
- NIH Public Access: Author ManuscriptDocument10 pagesNIH Public Access: Author ManuscriptErdelyi-Molnár ImolaNo ratings yet
- Udalov-Benkovskaya2011 Article PopulationGeneticsOfTheColoradDocument13 pagesUdalov-Benkovskaya2011 Article PopulationGeneticsOfTheColoradErdelyi-Molnár ImolaNo ratings yet
- GENOMIC DNA FROM INSECTSDocument2 pagesGENOMIC DNA FROM INSECTSErdelyi-Molnár ImolaNo ratings yet
- Detection of Organophosphate and Pyrethroid Resistance Alleles in Czech Colorado Potato Beetle PopulationsDocument8 pagesDetection of Organophosphate and Pyrethroid Resistance Alleles in Czech Colorado Potato Beetle PopulationsErdelyi-Molnár ImolaNo ratings yet
- BenKovskaya2008 Article TheGeneticBaseAndPhenotypicManDocument6 pagesBenKovskaya2008 Article TheGeneticBaseAndPhenotypicManErdelyi-Molnár ImolaNo ratings yet
- Udalov-Benkovskaya2011 Article ChangeInThePolymorphismLevelInDocument6 pagesUdalov-Benkovskaya2011 Article ChangeInThePolymorphismLevelInErdelyi-Molnár ImolaNo ratings yet
- Treatment of Acute BronchitisDocument3 pagesTreatment of Acute BronchitisjokosudibyoNo ratings yet
- Experimental and Correlational Studies of The Fear of Death: Direct Inethdds. The Earliest TechniqueDocument10 pagesExperimental and Correlational Studies of The Fear of Death: Direct Inethdds. The Earliest TechniqueDana PascuNo ratings yet
- BOSH For SO1 Student Manual SampleDocument65 pagesBOSH For SO1 Student Manual SampleFroilan olazo100% (1)
- A Survey On Data Mining Techniques For COVID PredictionDocument6 pagesA Survey On Data Mining Techniques For COVID PredictionWARSE Journals100% (2)
- DT Asa Bella-1Document12 pagesDT Asa Bella-1Bella Faradiska YuandaNo ratings yet
- Addressing Barriers, Stigma and Discrimination Surrounding HIV and Viral HepatitisDocument17 pagesAddressing Barriers, Stigma and Discrimination Surrounding HIV and Viral HepatitisAlemayehu KebedeNo ratings yet
- OVPR FY20Q1 Proposal List PDFDocument45 pagesOVPR FY20Q1 Proposal List PDFtsuregiNo ratings yet
- Republic of The Philippines Department of Education Region V - BicolDocument2 pagesRepublic of The Philippines Department of Education Region V - BicolKevin Viernes GaraisNo ratings yet
- Nilufa Shariff - Pain Dissertation PresentationDocument31 pagesNilufa Shariff - Pain Dissertation PresentationNilufar JivrajNo ratings yet
- Anti - HyperlipidemiaDocument18 pagesAnti - HyperlipidemiaZakarie Abdullahi Hussein100% (1)
- Telaah Kritis Artikel Terapi PZ 2020Document7 pagesTelaah Kritis Artikel Terapi PZ 2020dewi arifahniNo ratings yet
- Effect of Deep Breathing on Blood Pressure in Hypertensive PatientsDocument10 pagesEffect of Deep Breathing on Blood Pressure in Hypertensive PatientssendyNo ratings yet
- 07 - Towards Elimination DR Gottfried HirnschallDocument24 pages07 - Towards Elimination DR Gottfried HirnschallAjeet LohanaNo ratings yet
- Baby Thesis RevisedDocument20 pagesBaby Thesis RevisedSanjoe Angelo ManaloNo ratings yet
- Neet Ug Absolute Biology Vol 2Document22 pagesNeet Ug Absolute Biology Vol 2Rakesh Agarwal0% (1)
- 1 PBDocument11 pages1 PBNaswa Alifia PutriNo ratings yet
- Estate of Gerardo Gutierrez vs. PublixDocument12 pagesEstate of Gerardo Gutierrez vs. PublixPeterBurkeNo ratings yet
- International Journal of Cardiology May 2021 Issue 1Document275 pagesInternational Journal of Cardiology May 2021 Issue 1Fareesha KhanNo ratings yet
- Traditional Zootherapeutic Uses in The Treatment of Asthma by The Ethnic Groups of Assam, IndiaDocument5 pagesTraditional Zootherapeutic Uses in The Treatment of Asthma by The Ethnic Groups of Assam, IndiaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Cataract Qustion and AnswersDocument90 pagesCataract Qustion and AnswersVaishnavi MNo ratings yet
- AVBS1003-2020-Lecture 3-Anthrozoology IDocument25 pagesAVBS1003-2020-Lecture 3-Anthrozoology IOttilia LaiNo ratings yet
- Info - PathophysiologyDocument6 pagesInfo - PathophysiologyRupert BassigNo ratings yet
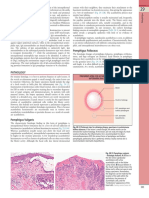
- Pemphigus Foliaceus: Fig. 29.13BDocument1 pagePemphigus Foliaceus: Fig. 29.13BCristian QuitoNo ratings yet
- Lista Detaliata A Serviciilor Regina MariaDocument69 pagesLista Detaliata A Serviciilor Regina MariaAmreusit SaschimbnumeleNo ratings yet
- Eat Yourself Smart: Britain Faces Airlift DeadlineDocument64 pagesEat Yourself Smart: Britain Faces Airlift DeadlineNidhi BhartiNo ratings yet
- Research Paper On Bells PalsyDocument5 pagesResearch Paper On Bells Palsyklbndecnd100% (1)
- Case Presentation: Presenter: DR Amanda Lundah Date: 15/4/20Document25 pagesCase Presentation: Presenter: DR Amanda Lundah Date: 15/4/20King MazingaNo ratings yet
- Community Question Bank (N.a)Document41 pagesCommunity Question Bank (N.a)Sumaya AfifyNo ratings yet
- Procedure Checklist For Suctioning NCM 112 SL Done Not DoneDocument4 pagesProcedure Checklist For Suctioning NCM 112 SL Done Not DoneIrish Eunice Felix100% (1)
- Specialized Group Homes For Persons With Severe or Profound Mental Retardation and Serious Problem Behaviour in EnglandDocument18 pagesSpecialized Group Homes For Persons With Severe or Profound Mental Retardation and Serious Problem Behaviour in EnglandDeborah RiskinNo ratings yet